

ATG2 transports lipids to promote autophagosome biogenesis
- PMID: 30952800
- PMCID: PMC6548141
- DOI: 10.1083/jcb.201811139
ATG2 transports lipids to promote autophagosome biogenesis
Abstract
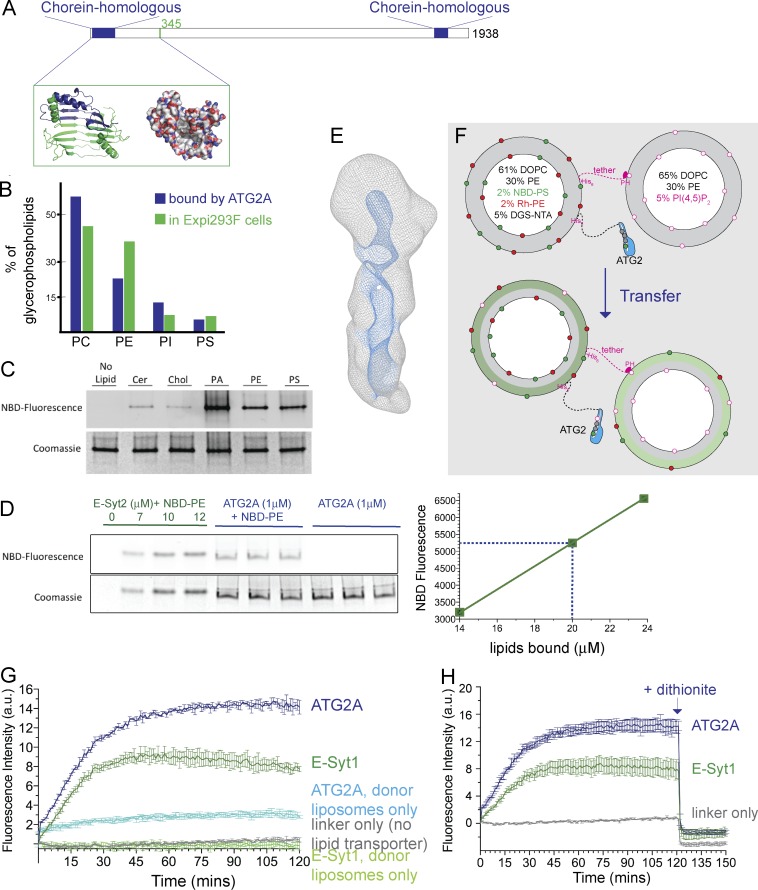
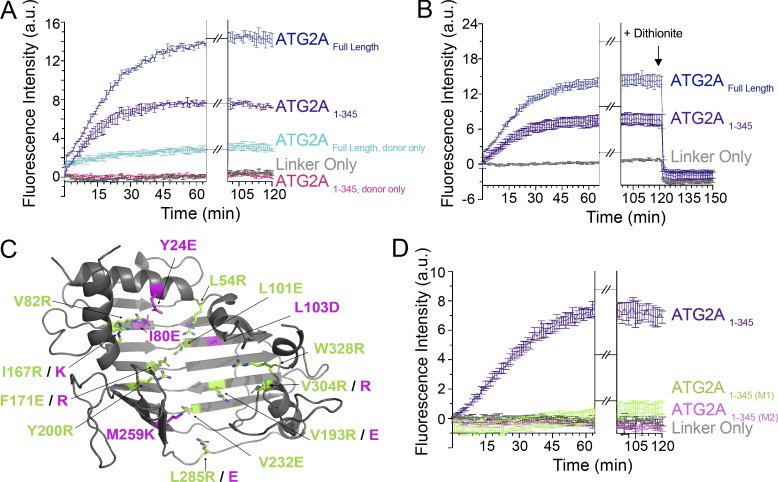
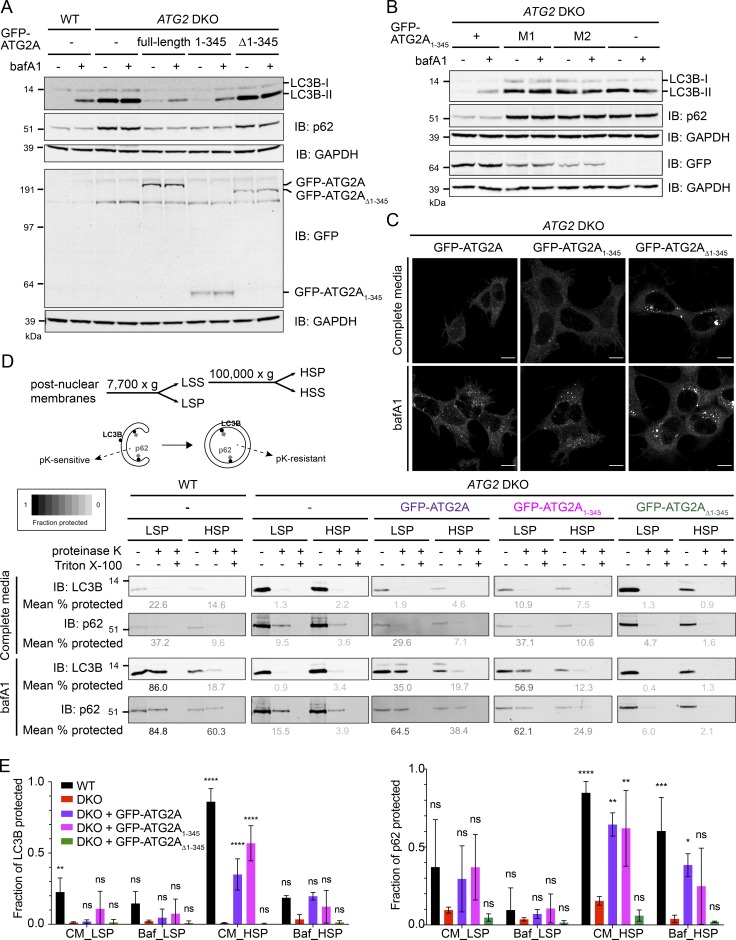
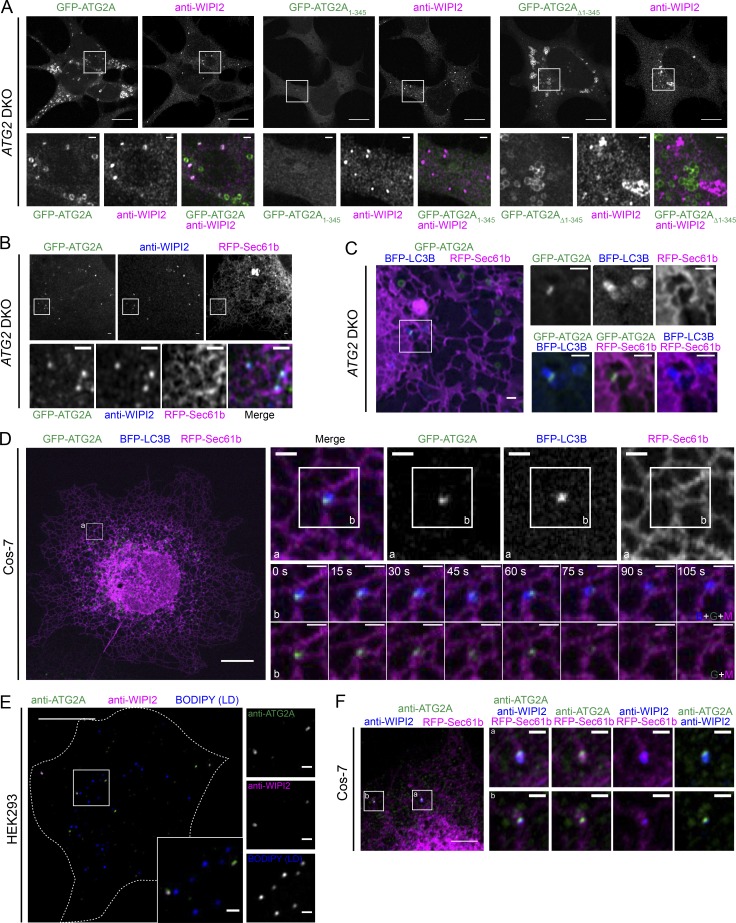
During macroautophagic stress, autophagosomes can be produced continuously and in high numbers. Many different organelles have been reported as potential donor membranes for this sustained autophagosome growth, but specific machinery to support the delivery of lipid to the growing autophagosome membrane has remained unknown. Here we show that the autophagy protein, ATG2, without a clear function since its discovery over 20 yr ago, is in fact a lipid-transfer protein likely operating at the ER-autophagosome interface. ATG2A can bind tens of glycerophospholipids at once and transfers lipids robustly in vitro. An N-terminal fragment of ATG2A that supports lipid transfer in vitro is both necessary and fully sufficient to rescue blocked autophagosome biogenesis in ATG2A/ATG2B KO cells, implying that regulation of lipid homeostasis is the major autophagy-dependent activity of this protein and, by extension, that protein-mediated lipid transfer across contact sites is a principal contributor to autophagosome formation.
© 2019 Valverde et al.
Figures
Comment in
-
Who plays the ferryman: ATG2 channels lipids into the forming autophagosome.J Cell Biol. 2019 Jun 3;218(6):1767-1768. doi: 10.1083/jcb.201904159. Epub 2019 May 10. J Cell Biol. 2019. PMID: 31076453 Free PMC article.
References
-
- Axe E.L., Walker S.A., Manifava M., Chandra P., Roderick H.L., Habermann A., Griffiths G., and Ktistakis N.T.. 2008. Autophagosome formation from membrane compartments enriched in phosphatidylinositol 3-phosphate and dynamically connected to the endoplasmic reticulum. J. Cell Biol. 182:685–701. 10.1083/jcb.200803137 - DOI - PMC - PubMed
-
- Gómez-Sánchez R., Rose J., Guimarães R., Mari M., Papinski D., Rieter E., Geerts W.J., Hardenberg R., Kraft C., Ungermann C., and Reggiori F.. 2018. Atg9 establishes Atg2-dependent contact sites between the endoplasmic reticulum and phagophores. J. Cell Biol. 217:2743–2763. 10.1083/jcb.201710116 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
