

A revised biosynthetic pathway for the cofactor F420 in prokaryotes
- PMID: 30952857
- PMCID: PMC6450877
- DOI: 10.1038/s41467-019-09534-x
A revised biosynthetic pathway for the cofactor F420 in prokaryotes
Abstract
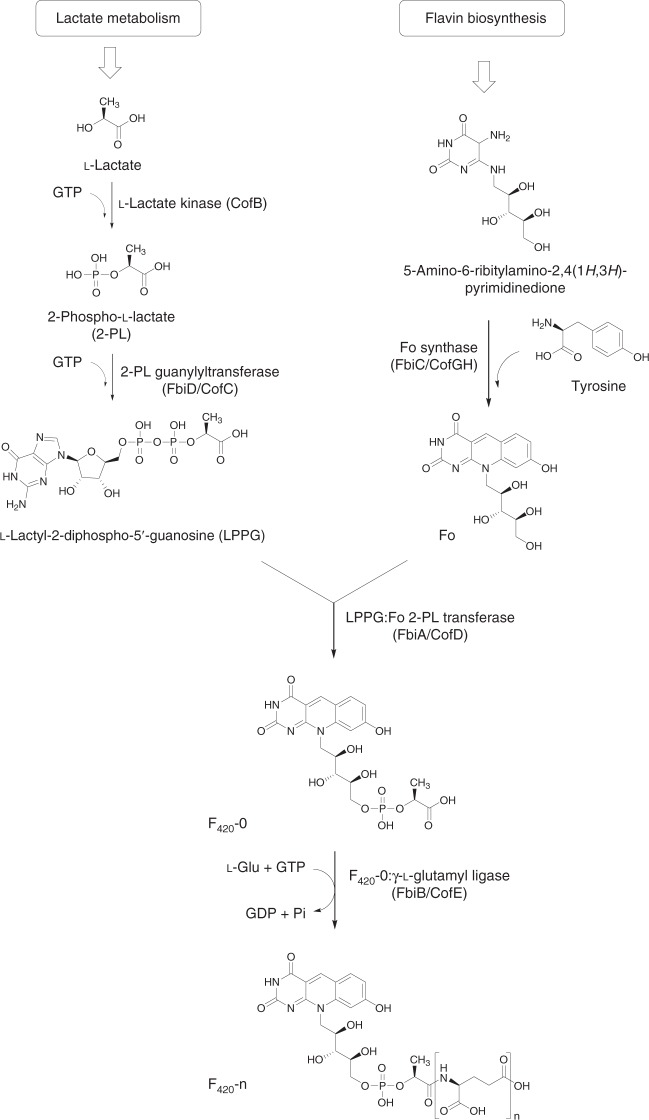
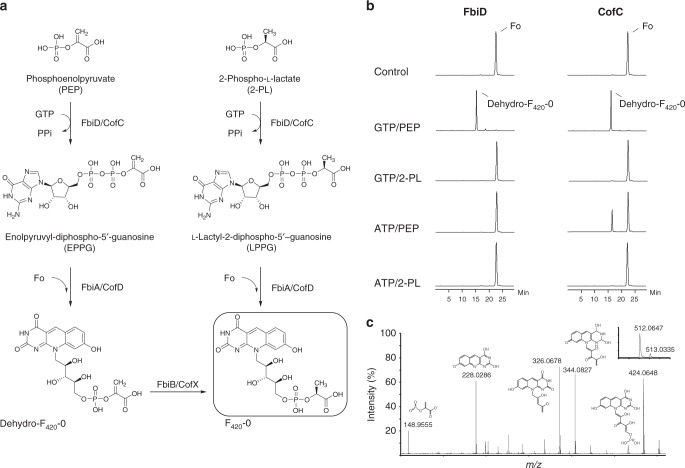
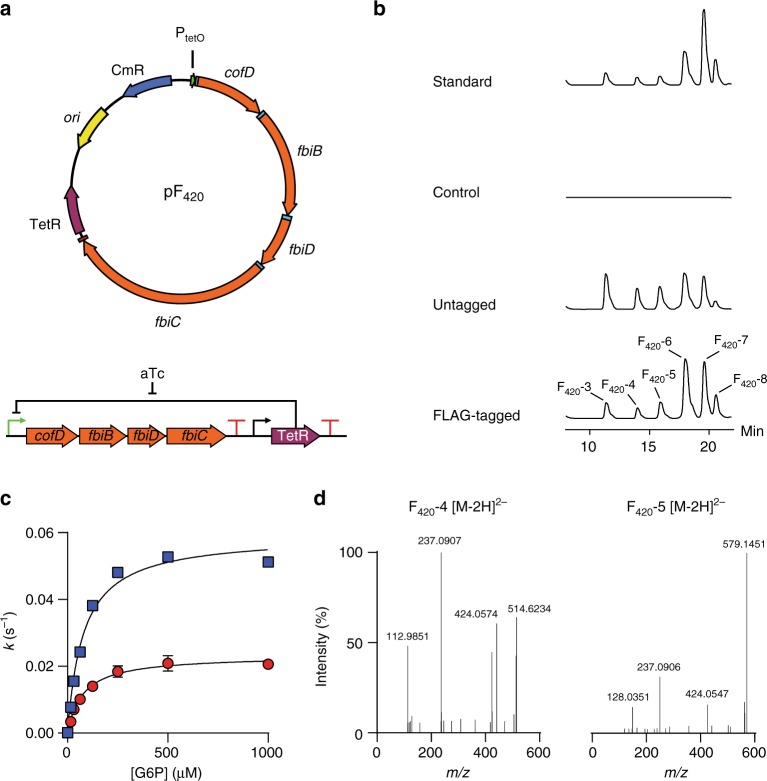
Cofactor F420 plays critical roles in primary and secondary metabolism in a range of bacteria and archaea as a low-potential hydride transfer agent. It mediates a variety of important redox transformations involved in bacterial persistence, antibiotic biosynthesis, pro-drug activation and methanogenesis. However, the biosynthetic pathway for F420 has not been fully elucidated: neither the enzyme that generates the putative intermediate 2-phospho-L-lactate, nor the function of the FMN-binding C-terminal domain of the γ-glutamyl ligase (FbiB) in bacteria are known. Here we present the structure of the guanylyltransferase FbiD and show that, along with its archaeal homolog CofC, it accepts phosphoenolpyruvate, rather than 2-phospho-L-lactate, as the substrate, leading to the formation of the previously uncharacterized intermediate dehydro-F420-0. The C-terminal domain of FbiB then utilizes FMNH2 to reduce dehydro-F420-0, which produces mature F420 species when combined with the γ-glutamyl ligase activity of the N-terminal domain. These new insights have allowed the heterologous production of F420 from a recombinant F420 biosynthetic pathway in Escherichia coli.
Conflict of interest statement
The authors declare no competing interests.
Figures



References
-
- Walsh C. Naturally occurring 5-deazaflavin coenzymes: biological redox roles. Acc. Chem. Res. 1986;19:216–221. doi: 10.1021/ar00127a004. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
