

BEAST 2.5: An advanced software platform for Bayesian evolutionary analysis
- PMID: 30958812
- PMCID: PMC6472827
- DOI: 10.1371/journal.pcbi.1006650
BEAST 2.5: An advanced software platform for Bayesian evolutionary analysis
Abstract
Elaboration of Bayesian phylogenetic inference methods has continued at pace in recent years with major new advances in nearly all aspects of the joint modelling of evolutionary data. It is increasingly appreciated that some evolutionary questions can only be adequately answered by combining evidence from multiple independent sources of data, including genome sequences, sampling dates, phenotypic data, radiocarbon dates, fossil occurrences, and biogeographic range information among others. Including all relevant data into a single joint model is very challenging both conceptually and computationally. Advanced computational software packages that allow robust development of compatible (sub-)models which can be composed into a full model hierarchy have played a key role in these developments. Developing such software frameworks is increasingly a major scientific activity in its own right, and comes with specific challenges, from practical software design, development and engineering challenges to statistical and conceptual modelling challenges. BEAST 2 is one such computational software platform, and was first announced over 4 years ago. Here we describe a series of major new developments in the BEAST 2 core platform and model hierarchy that have occurred since the first release of the software, culminating in the recent 2.5 release.
Conflict of interest statement
The authors have declared that no competing interests exist.
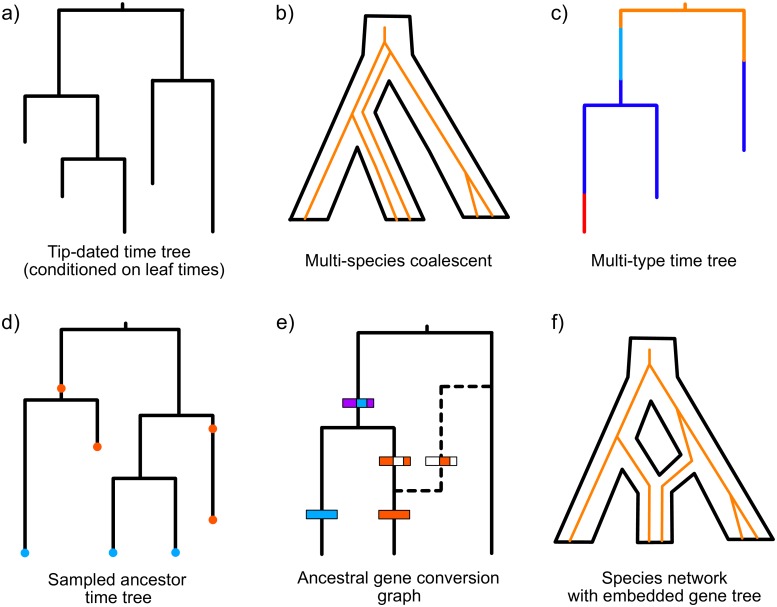
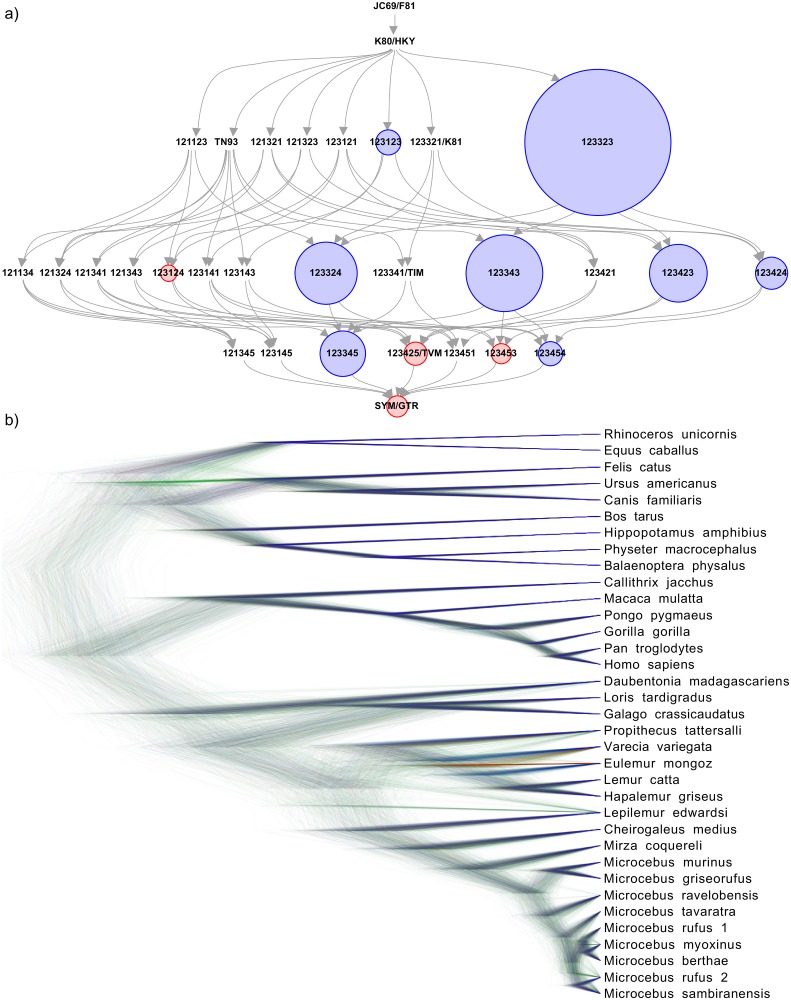
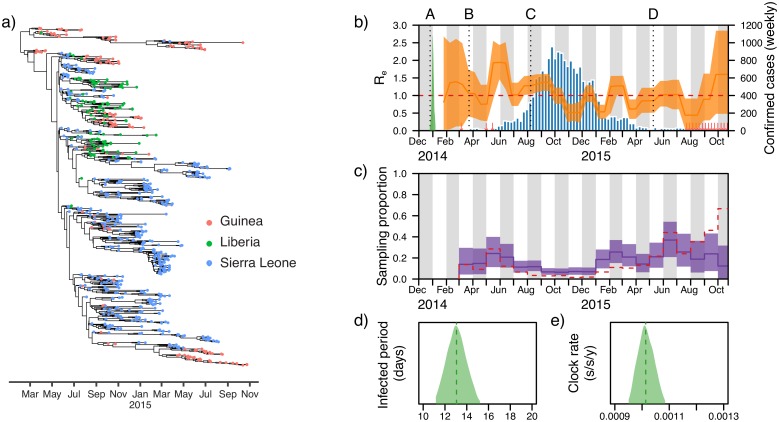
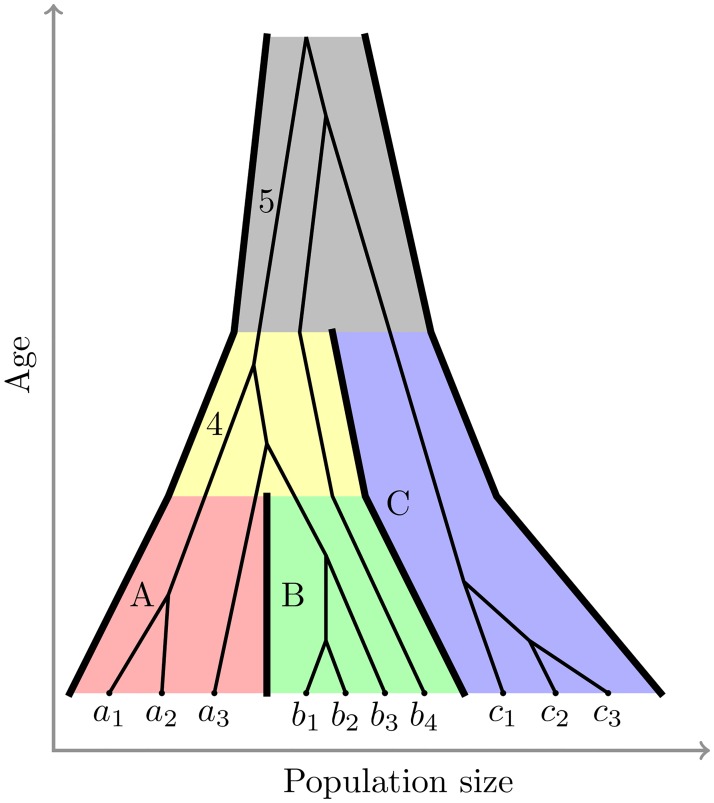
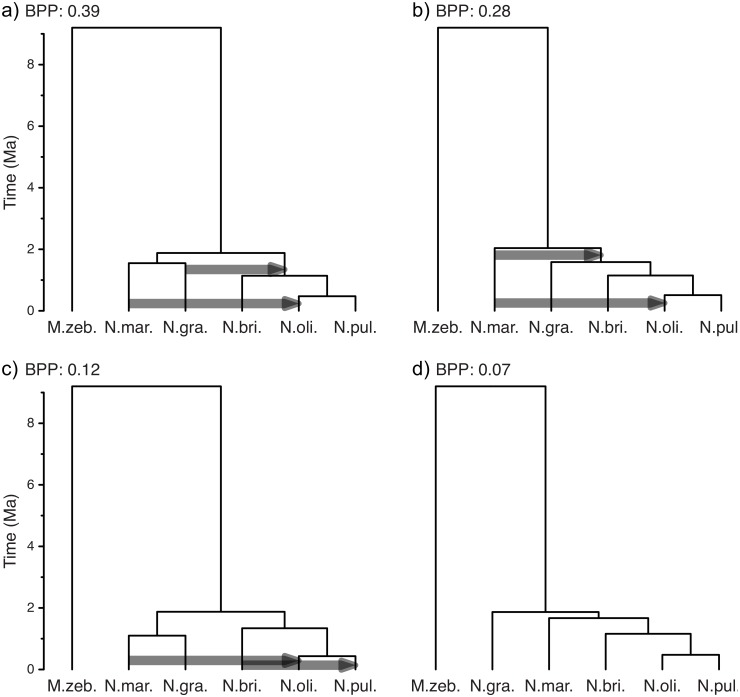
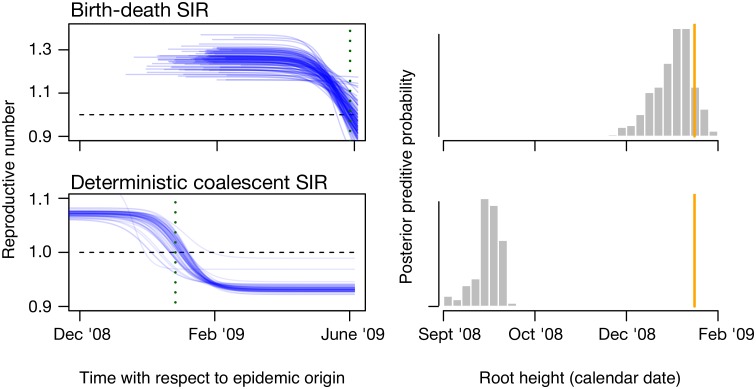
Figures
References
-
- Drummond AJ, Bouckaert RR. Bayesian evolutionary analysis with BEAST. Cambridge University Press; 2015.
-
- Bouckaert R, Heled J. DensiTree 2: Seeing trees through the forest. bioRxiv. 2014; p. 012401.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
