

Chromatin Loop Extrusion and Chromatin Unknotting
- PMID: 30961051
- PMCID: PMC6403842
- DOI: 10.3390/polym10101126
Chromatin Loop Extrusion and Chromatin Unknotting
Abstract
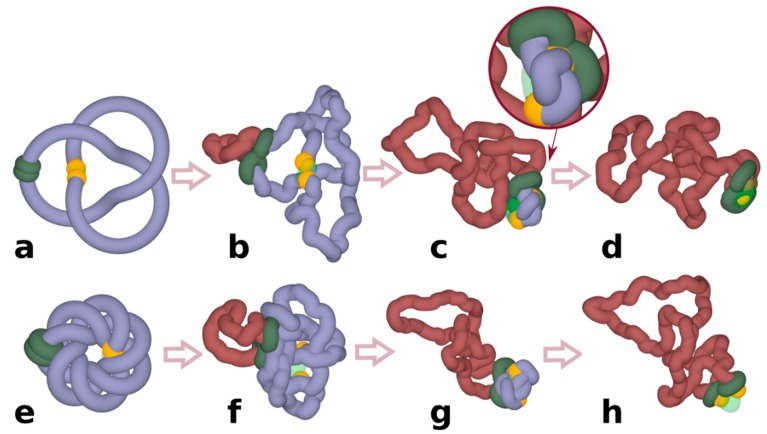
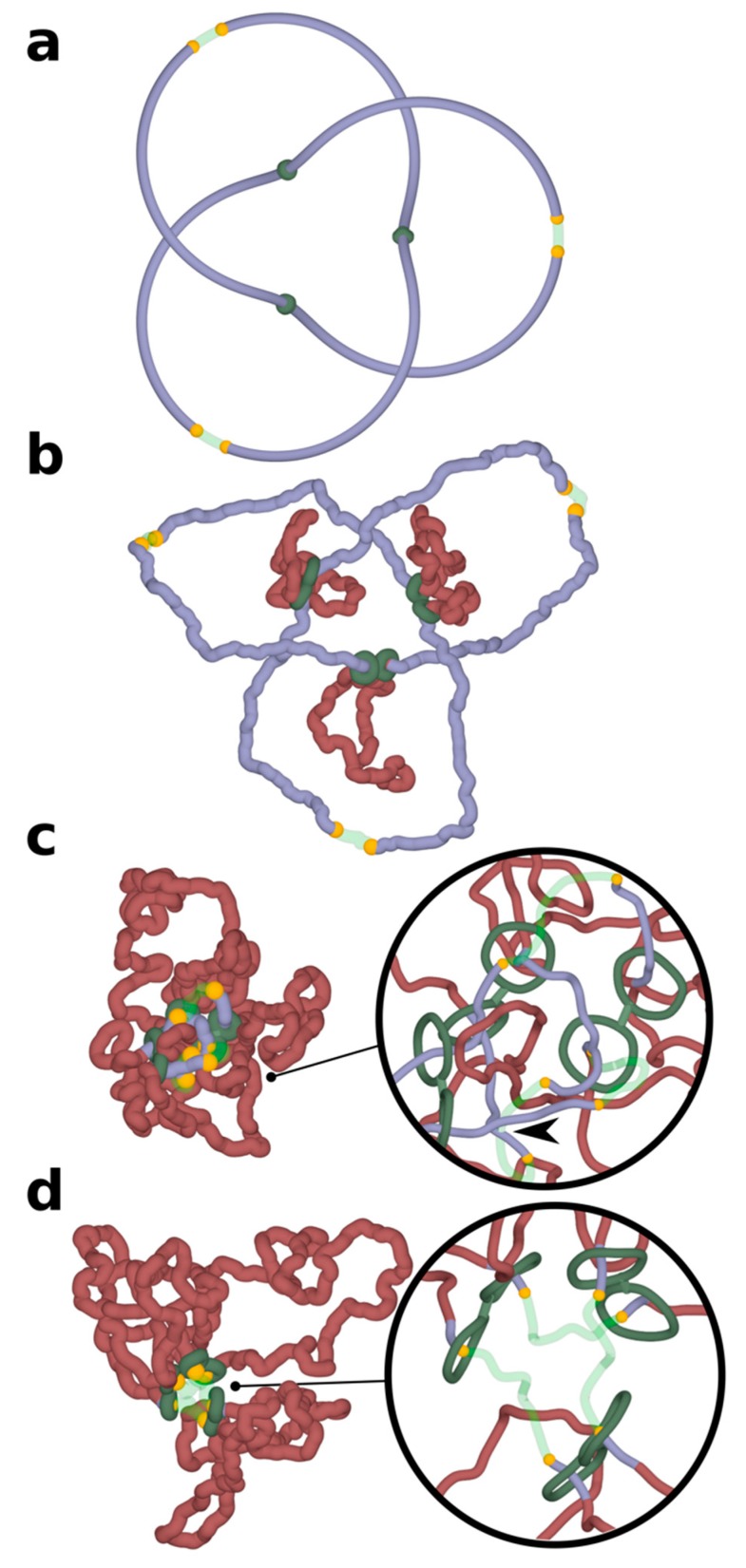
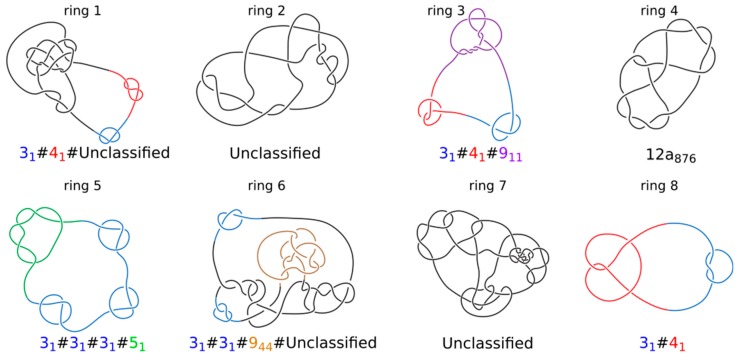
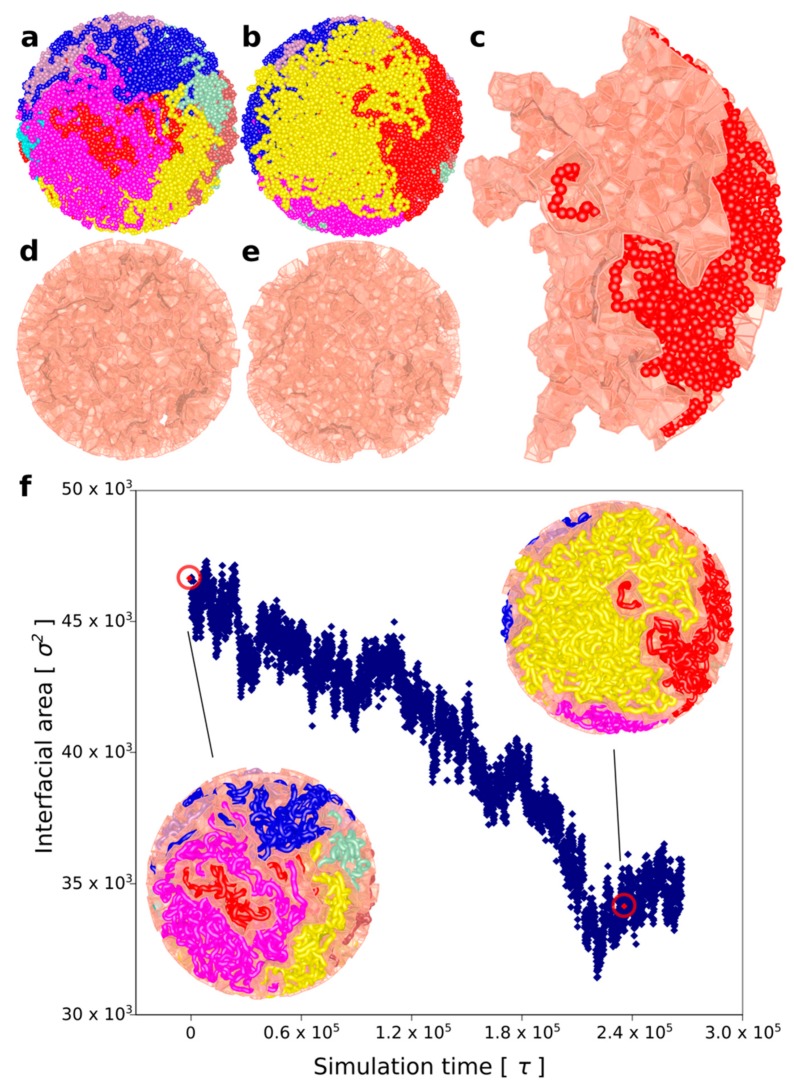
It has been a puzzle how decondensed interphase chromosomes remain essentially unknotted. The natural expectation is that in the presence of type II DNA topoisomerases that permit passages of double-stranded DNA regions through each other, all chromosomes should reach the state of topological equilibrium. The topological equilibrium in highly crowded interphase chromosomes forming chromosome territories would result in formation of highly knotted chromatin fibres. However, Chromosome Conformation Capture (3C) methods revealed that the decay of contact probabilities with the genomic distance in interphase chromosomes is practically the same as in the crumpled globule state that is formed when long polymers condense without formation of any knots. To remove knots from highly crowded chromatin, one would need an active process that should not only provide the energy to move the system from the state of topological equilibrium but also guide topoisomerase-mediated passages in such a way that knots would be efficiently unknotted instead of making the knots even more complex. We perform coarse-grained molecular dynamics simulations of the process of chromatin loop extrusion involving knotted and catenated chromatin fibres to check whether chromatin loop extrusion may be involved in active unknotting of chromatin fibres. Our simulations show that the process of chromatin loop extrusion is ideally suited to actively unknot, decatenate and demix chromatin fibres in interphase chromosomes.
Keywords: DNA knots; DNA topoisomerases; biopolymers; chromatin; chromatin loop extrusion; chromosomes; cohesin.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Figures
References
-
- Bates A.D., Maxwell A. DNA Topology. Oxford University Press; Oxford, UK: 2005. p. 198.
Grants and funding
LinkOut - more resources
Full Text Sources
