

ATAC-seq reveals regional differences in enhancer accessibility during the establishment of spatial coordinates in the Drosophila blastoderm
- PMID: 30962180
- PMCID: PMC6499308
- DOI: 10.1101/gr.242362.118
ATAC-seq reveals regional differences in enhancer accessibility during the establishment of spatial coordinates in the Drosophila blastoderm
Abstract
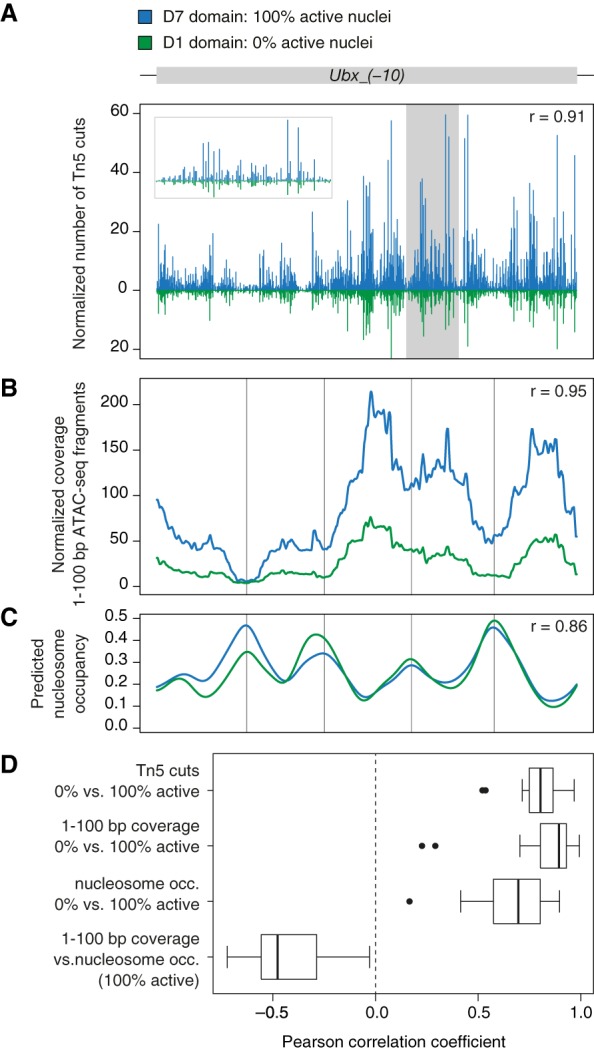
Establishment of spatial coordinates during Drosophila embryogenesis relies on differential regulatory activity of axis patterning enhancers. Concentration gradients of activator and repressor transcription factors (TFs) provide positional information to each enhancer, which in turn promotes transcription of a target gene in a specific spatial pattern. However, the interplay between an enhancer regulatory activity and its accessibility as determined by local chromatin organization is not well understood. We profiled chromatin accessibility with ATAC-seq in narrow, genetically tagged domains along the antero-posterior axis in the Drosophila blastoderm. We demonstrate that one-quarter of the accessible genome displays significant regional variation in its ATAC-seq signal immediately after zygotic genome activation. Axis patterning enhancers are enriched among the most variable intervals, and their accessibility changes correlate with their regulatory activity. In an embryonic domain where an enhancer receives a net activating TF input and promotes transcription, it displays elevated accessibility in comparison to a domain where it receives a net repressive input. We propose that differential accessibility is a signature of patterning cis-regulatory elements in the Drosophila blastoderm and discuss potential mechanisms by which accessibility of enhancers may be modulated by activator and repressor TFs.
© 2019 Bozek et al.; Published by Cold Spring Harbor Laboratory Press.
Figures






References
-
- Bradley RK, Li XY, Trapnell C, Davidson S, Pachter L, Chu HC, Tonkin LA, Biggin MD, Eisen MB. 2010. Binding site turnover produces pervasive quantitative changes in transcription factor binding between closely related Drosophila species. PLoS Biol 8: e1000343 10.1371/journal.pbio.1000343 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous