

Working memory revived in older adults by synchronizing rhythmic brain circuits
- PMID: 30962628
- PMCID: PMC6486414
- DOI: 10.1038/s41593-019-0371-x
Working memory revived in older adults by synchronizing rhythmic brain circuits
Abstract
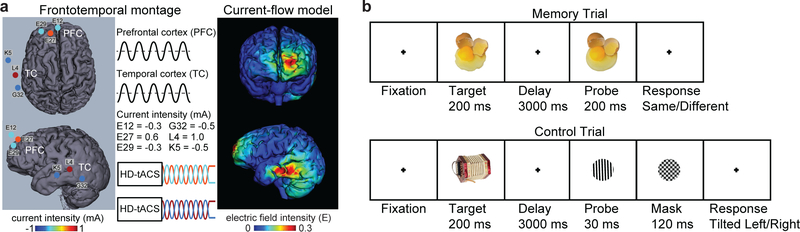
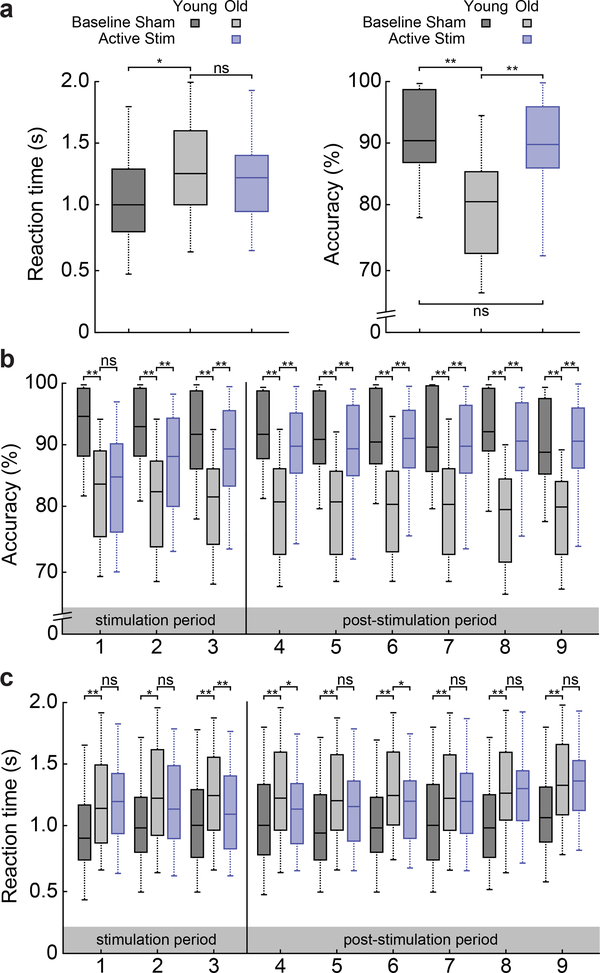
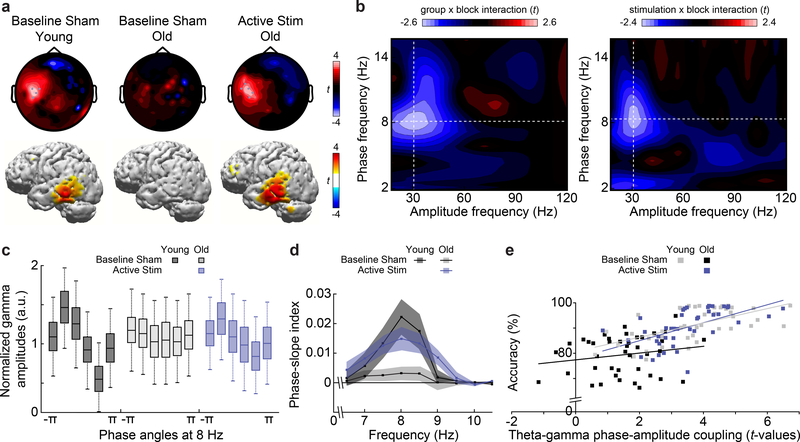
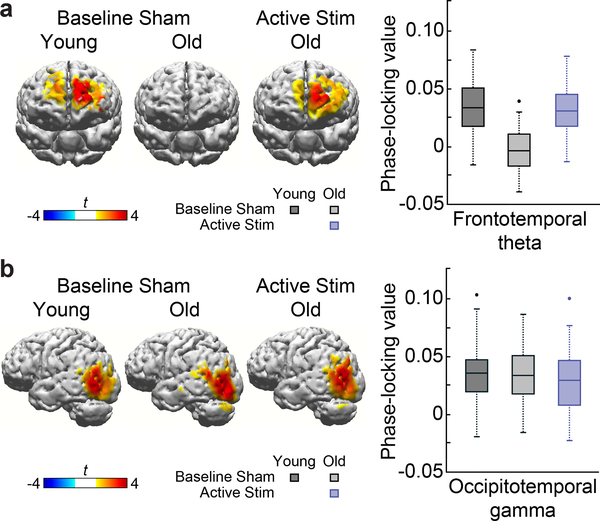
Understanding normal brain aging and developing methods to maintain or improve cognition in older adults are major goals of fundamental and translational neuroscience. Here we show a core feature of cognitive decline-working-memory deficits-emerges from disconnected local and long-range circuits instantiated by theta-gamma phase-amplitude coupling in temporal cortex and theta phase synchronization across frontotemporal cortex. We developed a noninvasive stimulation procedure for modulating long-range theta interactions in adults aged 60-76 years. After 25 min of stimulation, frequency-tuned to individual brain network dynamics, we observed a preferential increase in neural synchronization patterns and the return of sender-receiver relationships of information flow within and between frontotemporal regions. The end result was rapid improvement in working-memory performance that outlasted a 50 min post-stimulation period. The results provide insight into the physiological foundations of age-related cognitive impairment and contribute to groundwork for future non-pharmacological interventions targeting aspects of cognitive decline.
Conflict of interest statement
Competing interests
The authors declare no competing interests.
Figures
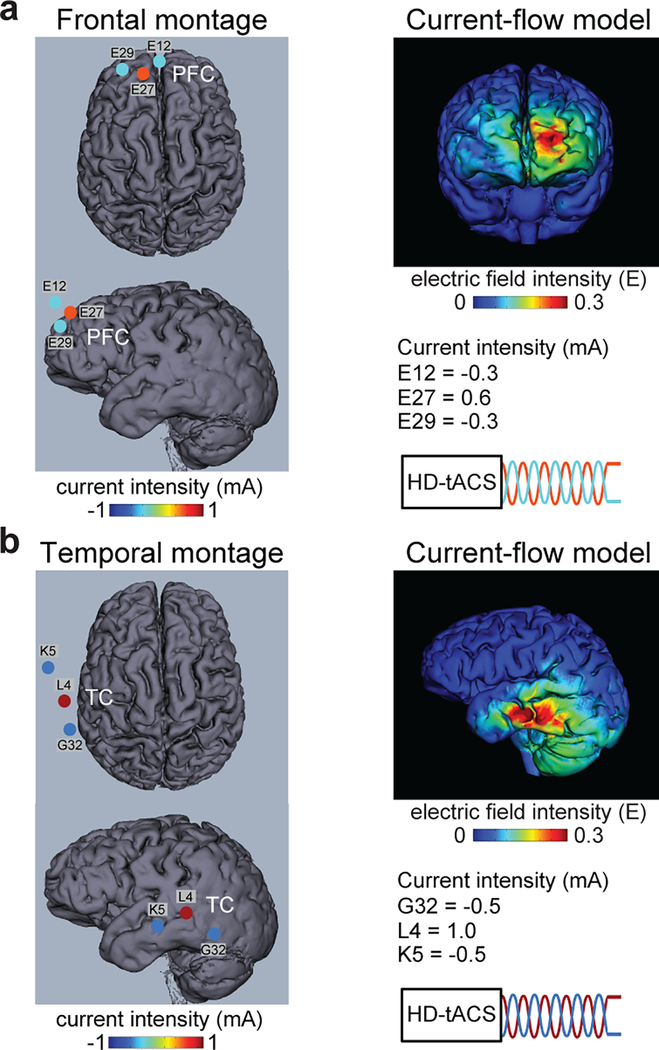
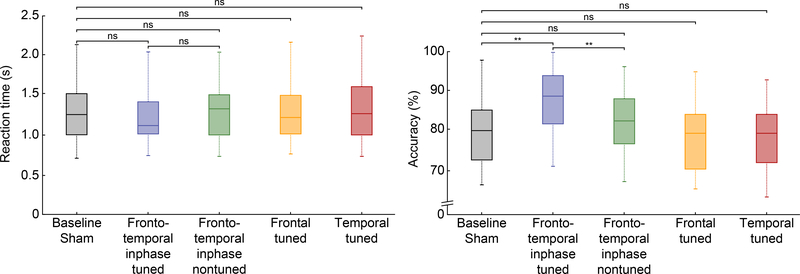
Comment in
-
Reversing working memory decline in the elderly.Nat Neurosci. 2019 May;22(5):686-688. doi: 10.1038/s41593-019-0386-3. Nat Neurosci. 2019. PMID: 30962629 No abstract available.
-
A memory boost for the elderly.Nat Hum Behav. 2019 Jun;3(6):547. doi: 10.1038/s41562-019-0608-4. Nat Hum Behav. 2019. PMID: 31048745 No abstract available.
References
-
- Hayutin AM Global demographic shifts create challenges and opportunities. PREA Quart, 46–53 (2007).
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
