

Interaction of viral pathogen with porin channels on the outer membrane of insect bacterial symbionts mediates their joint transovarial transmission
- PMID: 30967020
- PMCID: PMC6367154
- DOI: 10.1098/rstb.2018.0320
Interaction of viral pathogen with porin channels on the outer membrane of insect bacterial symbionts mediates their joint transovarial transmission
Abstract
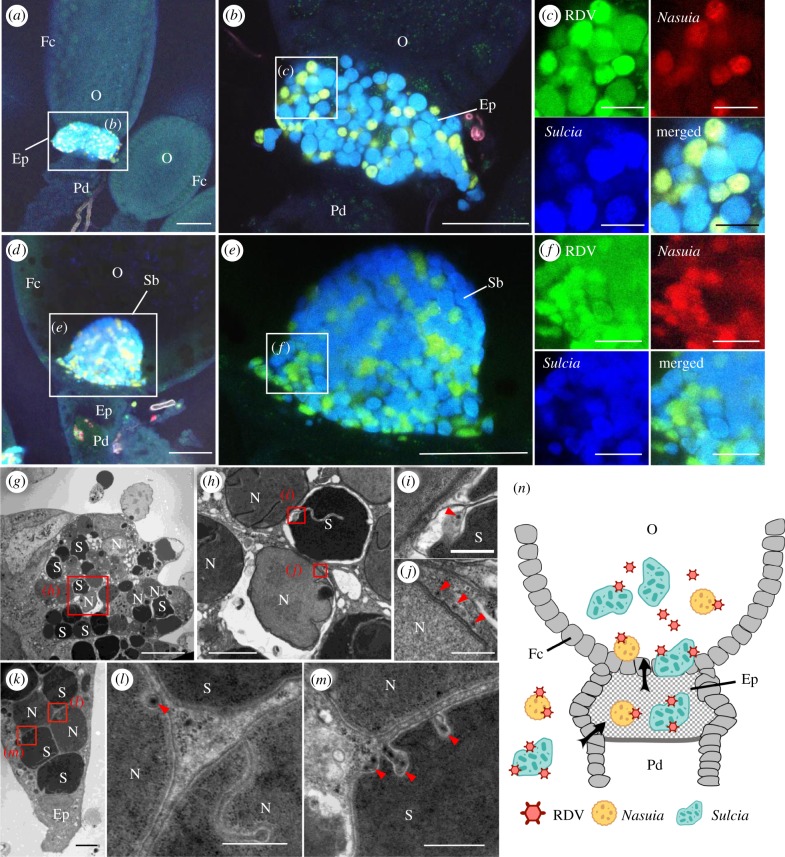
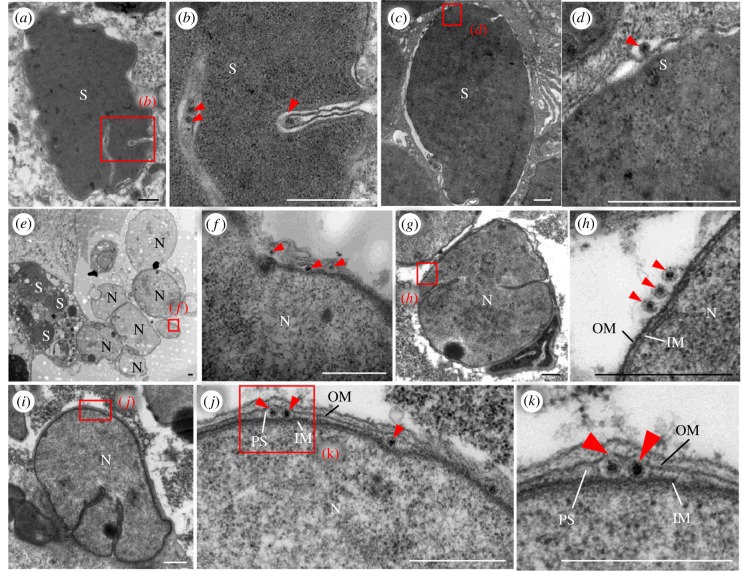
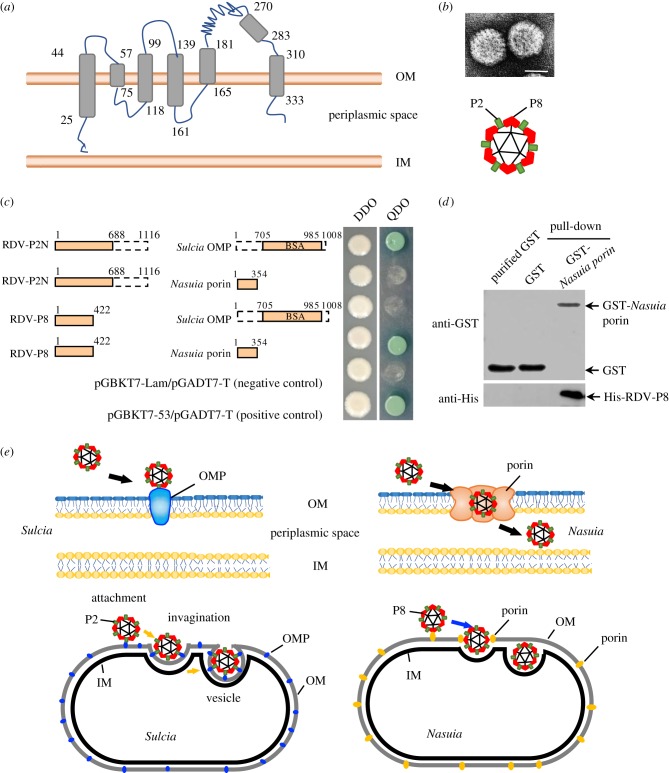
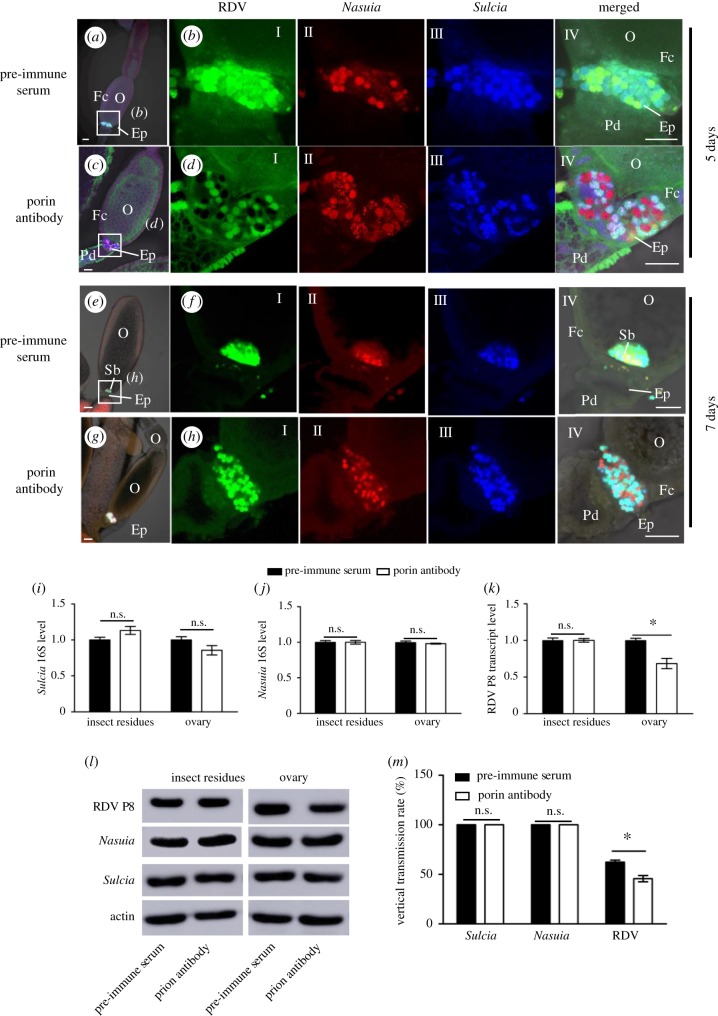
Many hemipteran insects that can transmit plant viruses in a persistent and transovarial manner are generally associated with a common obligate bacterial symbiont Sulcia and its β-proteobacterial partner. Rice dwarf virus (RDV), a plant reovirus, can bind to the envelope of Sulcia through direct interaction of the viral minor outer capsid protein P2 with the bacterial outer membrane protein, allowing the virus to exploit the ancient oocyte entry path of Sulcia in rice leafhopper vectors. Here, we show that RDV can hitchhike with both Sulcia and its β-proteobacterial partner Nasuia to ensure their simultaneous transovarial transmission. Interestingly, RDV can move through the outer envelope of Nasuia and reside in the periplasmic space, which is mediated by the specific interaction of the viral major outer capsid protein P8 and the porin channel on the bacterial outer envelope. Nasuia porin-specific antibody efficiently interferes with the binding between RDV and the Nasuia envelope, thus strongly preventing viral transmission to insect offspring. Thus, RDV has evolved different strategies to exploit the ancient oocyte entry paths used by two obligate bacterial symbionts in rice leafhoppers. Our results thus reveal that RDV has formed complex, cooperative interactions with both Sulcia and Nasuia during their joint transovarial transmission. This article is part of the theme issue 'Biotic signalling sheds light on smart pest management'.
Keywords: Nasuia and Sulcia; obligate bacterial symbionts; porin channel; rice dwarf virus; rice leafhopper; transovarial transmission.
Conflict of interest statement
We declare we have no competing interests.
Figures
Similar articles
-
Insect symbiotic bacteria harbour viral pathogens for transovarial transmission.Nat Microbiol. 2017 Mar 6;2:17025. doi: 10.1038/nmicrobiol.2017.25. Nat Microbiol. 2017. PMID: 28263320
-
Insect Bacterial Symbiont-Mediated Vitellogenin Uptake into Oocytes To Support Egg Development.mBio. 2020 Nov 10;11(6):e01142-20. doi: 10.1128/mBio.01142-20. mBio. 2020. PMID: 33172995 Free PMC article.
-
Comparative Genomics of the Dual-Obligate Symbionts from the Treehopper, Entylia carinata (Hemiptera: Membracidae), Provide Insight into the Origins and Evolution of an Ancient Symbiosis.Genome Biol Evol. 2017 Jun 1;9(6):1803-1815. doi: 10.1093/gbe/evx134. Genome Biol Evol. 2017. PMID: 28854637 Free PMC article.
-
Rice Reoviruses in Insect Vectors.Annu Rev Phytopathol. 2016 Aug 4;54:99-120. doi: 10.1146/annurev-phyto-080615-095900. Epub 2016 May 25. Annu Rev Phytopathol. 2016. PMID: 27296147 Review.
-
Transovarial Transmission of Symbionts in Insects.Results Probl Cell Differ. 2017;63:43-67. doi: 10.1007/978-3-319-60855-6_3. Results Probl Cell Differ. 2017. PMID: 28779313 Review.
Cited by
-
Insect ribosome-rescuer Pelo-Hbs1 complex on sperm surface mediates paternal arbovirus transmission.Nat Commun. 2024 Aug 9;15(1):6817. doi: 10.1038/s41467-024-51020-6. Nat Commun. 2024. PMID: 39122673 Free PMC article.
-
Autophagy mediates a direct synergistic interaction during co-transmission of two distinct arboviruses by insect vectors.Sci China Life Sci. 2023 Jul;66(7):1665-1681. doi: 10.1007/s11427-022-2228-y. Epub 2023 Mar 9. Sci China Life Sci. 2023. PMID: 36917406
-
Roles of Bacterial Symbionts in Transmission of Plant Virus by Hemipteran Vectors.Front Microbiol. 2022 Jan 27;13:805352. doi: 10.3389/fmicb.2022.805352. eCollection 2022. Front Microbiol. 2022. PMID: 35154053 Free PMC article. Review.
-
Overview: biotic signalling for smart pest management.Philos Trans R Soc Lond B Biol Sci. 2019 Mar 4;374(1767):20180306. doi: 10.1098/rstb.2018.0306. Philos Trans R Soc Lond B Biol Sci. 2019. PMID: 30967024 Free PMC article.
-
Single-cell RNA-seq reveals distinct metabolic "microniches" and close host-symbiont interactions in deep-sea chemosynthetic tubeworm.Sci Adv. 2024 Jul 26;10(30):eadn3053. doi: 10.1126/sciadv.adn3053. Epub 2024 Jul 24. Sci Adv. 2024. PMID: 39047091 Free PMC article.
References
Publication types
MeSH terms
Substances
Supplementary concepts
Associated data
LinkOut - more resources
Full Text Sources
