

Efficient allelic-drive in Drosophila
- PMID: 30967548
- PMCID: PMC6456580
- DOI: 10.1038/s41467-019-09694-w
Efficient allelic-drive in Drosophila
Abstract
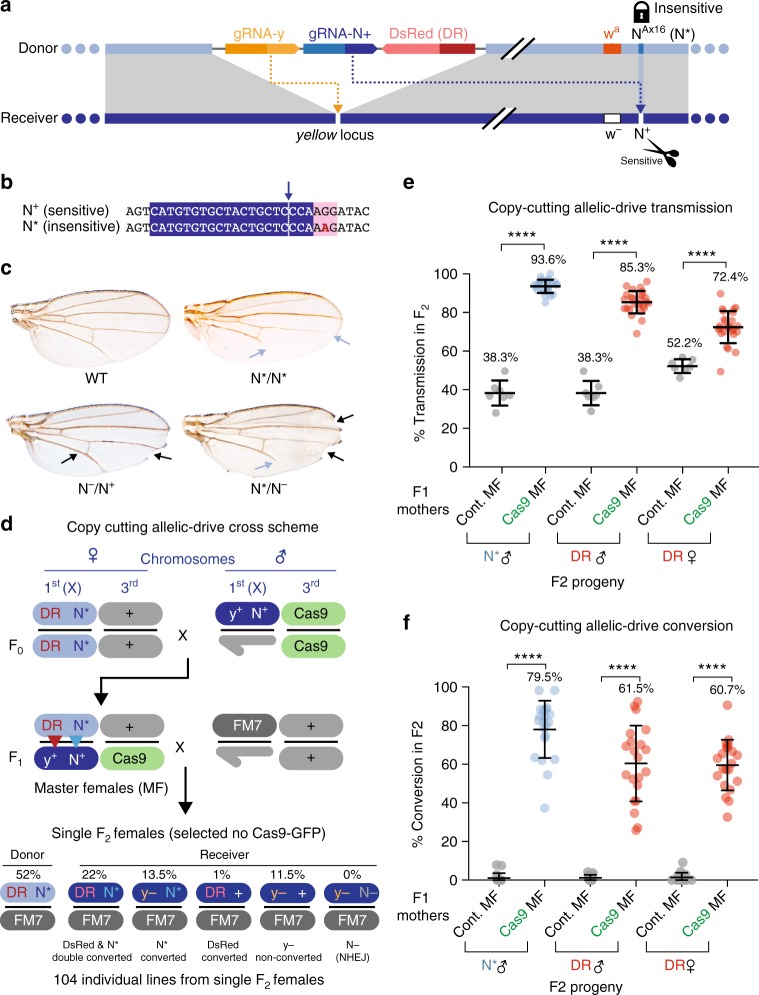
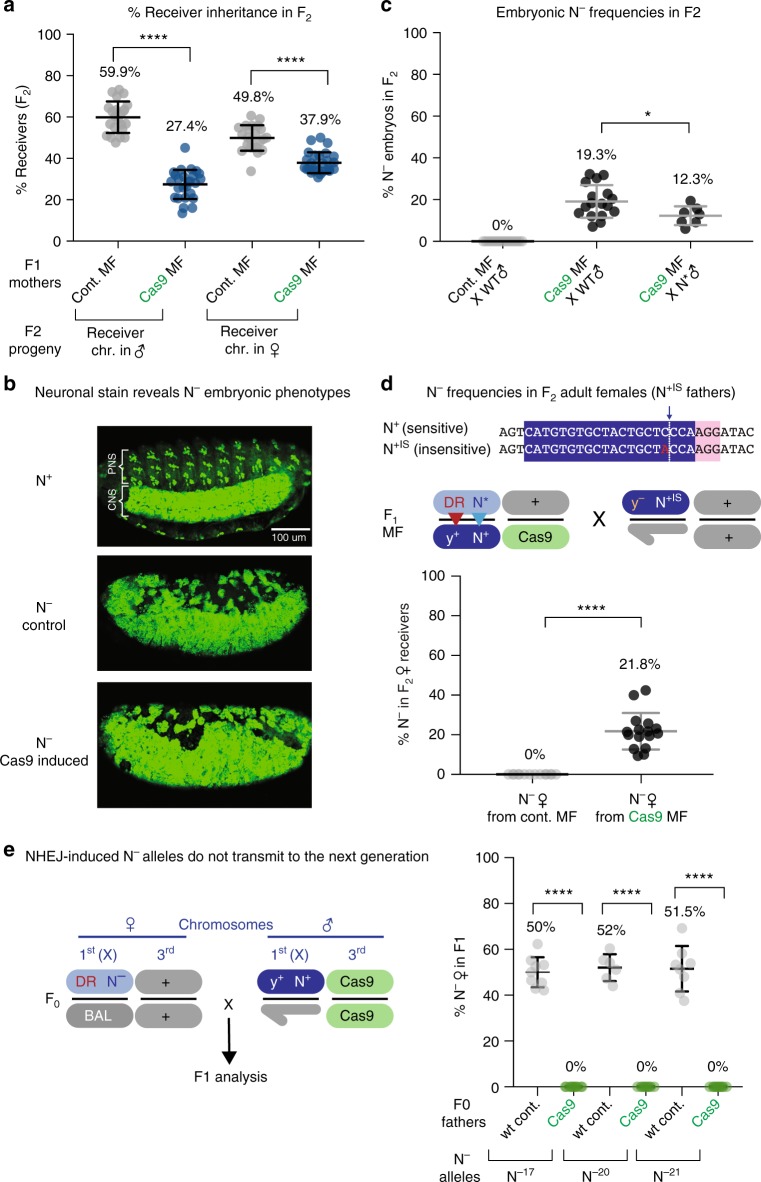
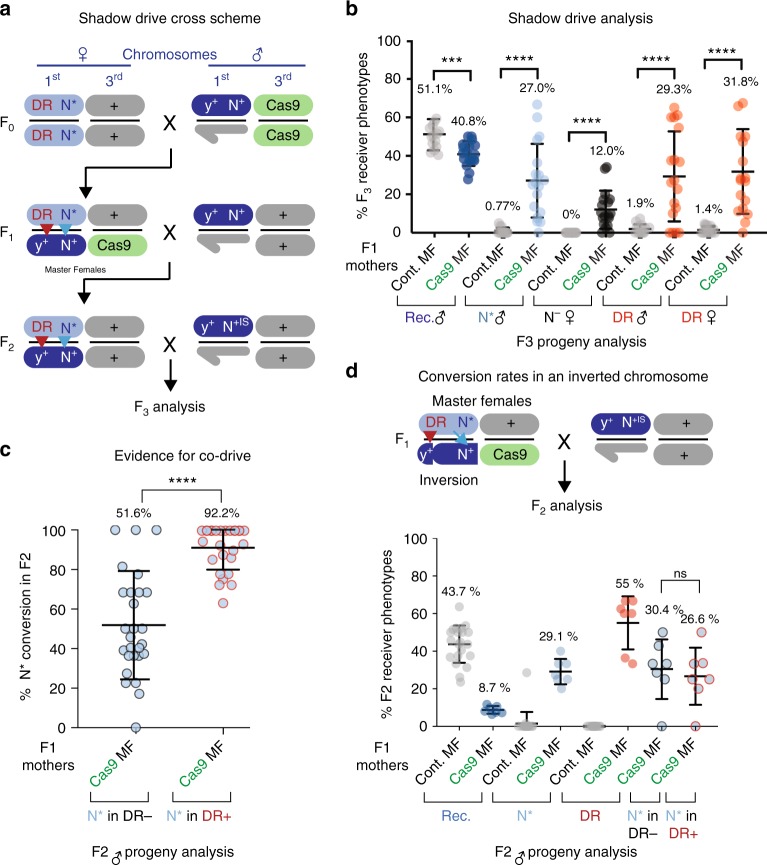
Gene-drive systems developed in several organisms result in super-Mendelian inheritance of transgenic insertions. Here, we generalize this "active genetic" approach to preferentially transmit allelic variants (allelic-drive) resulting from only a single or a few nucleotide alterations. We test two configurations for allelic-drive: one, copy-cutting, in which a non-preferred allele is selectively targeted for Cas9/guide RNA (gRNA) cleavage, and a more general approach, copy-grafting, that permits selective inheritance of a desired allele located in close proximity to the gRNA cut site. We also characterize a phenomenon we refer to as lethal-mosaicism that dominantly eliminates NHEJ-induced mutations and favors inheritance of functional cleavage-resistant alleles. These two efficient allelic-drive methods, enhanced by lethal mosaicism and a trans-generational drive process we refer to as "shadow-drive", have broad practical applications in improving health and agriculture and greatly extend the active genetics toolbox.
Conflict of interest statement
E.B. and V.G. have equity interest in Synbal Inc. and Agragene, Inc., companies that may potentially benefit from the research results and also serves on the company’s Board of Directors and Scientific Advisory Board. The terms of this arrangement have been reviewed and approved by the University of California, San Diego in accordance with its conflict of interest policies. The other authors declare no competing interests.
Figures

References
-
- DiCarlo, J. E., Chavez, A., Dietz, S. L., Esvelt, K. M. & Church, G. M. RNA-guided gene drives can efficiently bias inheritance in wild yeast. bioRxiv10.1101/013896 (2015).
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
