

Repeated Sigma-1 Receptor Antagonist MR309 Administration Modulates Central Neuropathic Pain Development After Spinal Cord Injury in Mice
- PMID: 30967775
- PMCID: PMC6439356
- DOI: 10.3389/fphar.2019.00222
Repeated Sigma-1 Receptor Antagonist MR309 Administration Modulates Central Neuropathic Pain Development After Spinal Cord Injury in Mice
Abstract
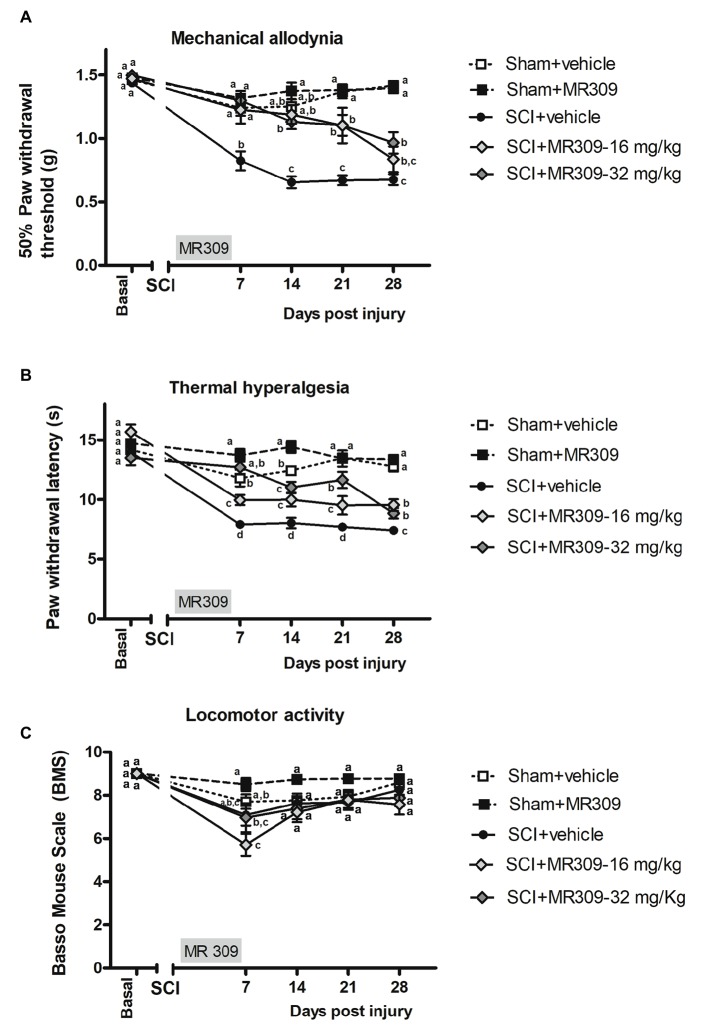
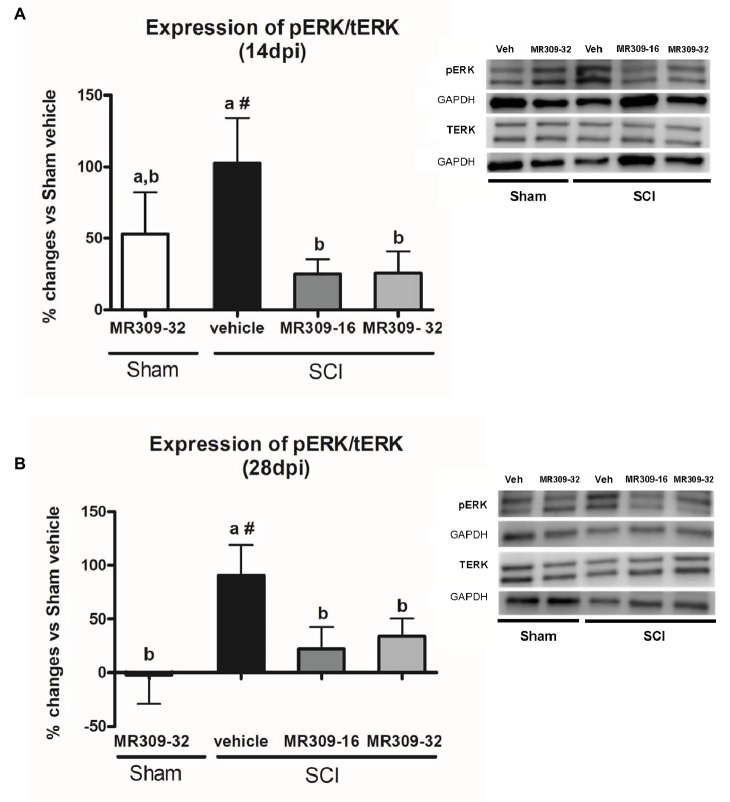
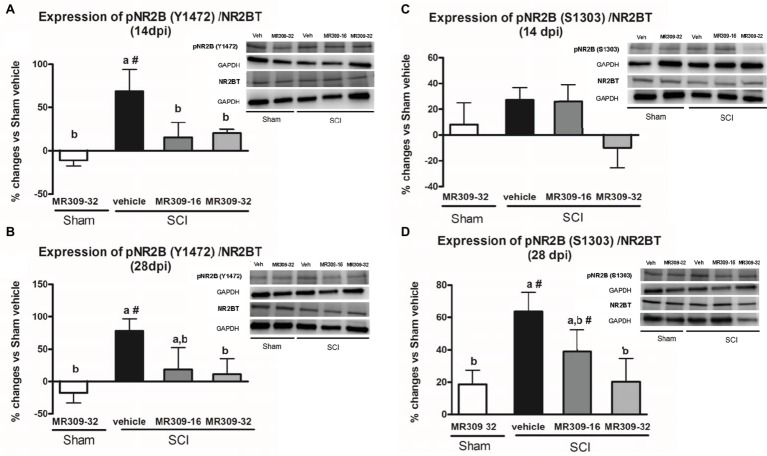
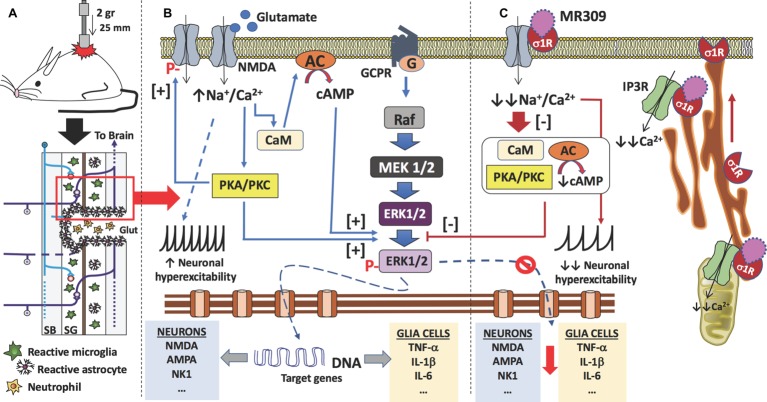
Up to two-thirds of patients affected by spinal cord injury (SCI) develop central neuropathic pain (CNP), which has a high impact on their quality of life. Most of the patients are largely refractory to current treatments, and new pharmacological strategies are needed. Recently, it has been shown that the acute administration of the σ1R antagonist MR309 (previously developed as E-52862) at 28 days after spinal cord contusion results in a dose-dependent suppression of both mechanical allodynia and thermal hyperalgesia in wild-type CD-1 Swiss female mice. The present work was addressed to determine whether MR309 might exert preventive effects on CNP development by repeated administration during the first week after SCI in mice. To this end, the MR309 (16 or 32 mg/kg i.p.) modulation on both thermal hyperalgesia and mechanical allodynia development were evaluated weekly up to 28 days post-injury. In addition, changes in pro-inflammatory cytokine (TNF-α, IL-1β) expression and both the expression and activation (phosphorylation) of the N-methyl-D-aspartate receptor subunit 2B (NR2B-NMDA) and extracellular signal-regulated kinases (ERK1/2) were analyzed. The repeated treatment of SCI-mice with MR309 resulted in significant pain behavior attenuation beyond the end of the administration period, accompanied by reduced expression of central sensitization-related mechanistic correlates, including extracellular mediators (TNF-α and IL-1β), membrane receptors/channels (NR2B-NMDA) and intracellular signaling cascades (ERK/pERK). These findings suggest that repeated MR309 treatment after SCI may be a suitable pharmacologic strategy to modulate SCI-induced CNP development.
Keywords: MR309; central neuropathic pain; central sensitization-related biomarkers; pro-inflammatory cytokines; spinal cord injury.
Figures

References
LinkOut - more resources
Full Text Sources
Miscellaneous