

Burkholderia pseudomallei BimC Is Required for Actin-Based Motility, Intracellular Survival, and Virulence
- PMID: 30968000
- PMCID: PMC6439308
- DOI: 10.3389/fcimb.2019.00063
Burkholderia pseudomallei BimC Is Required for Actin-Based Motility, Intracellular Survival, and Virulence
Abstract
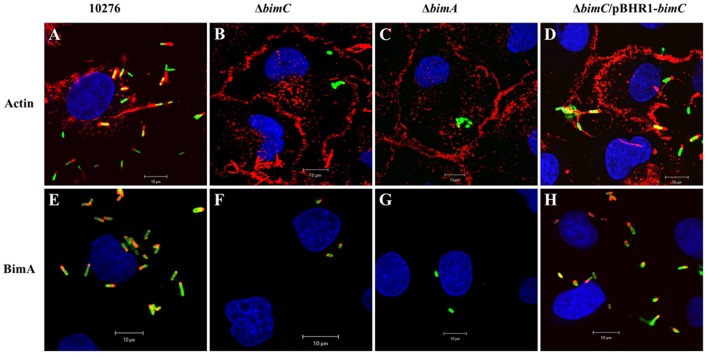
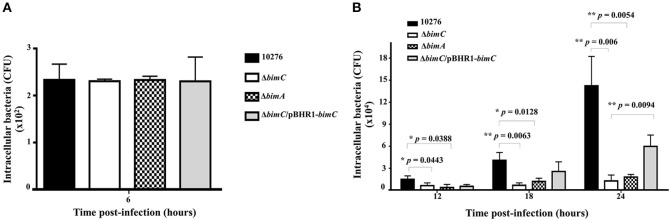
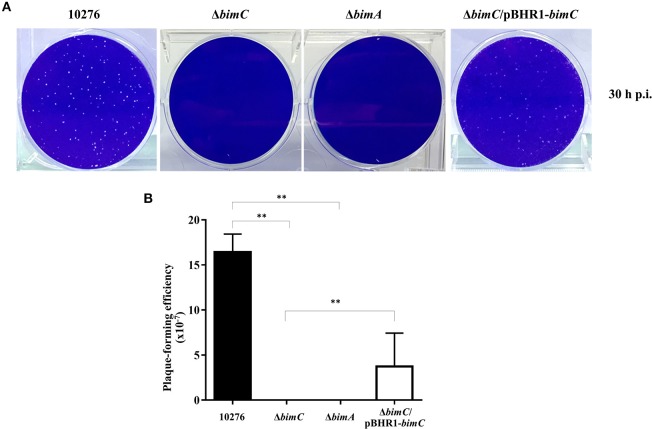
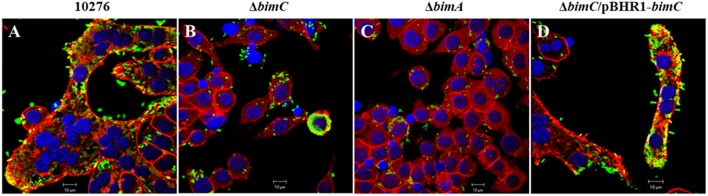
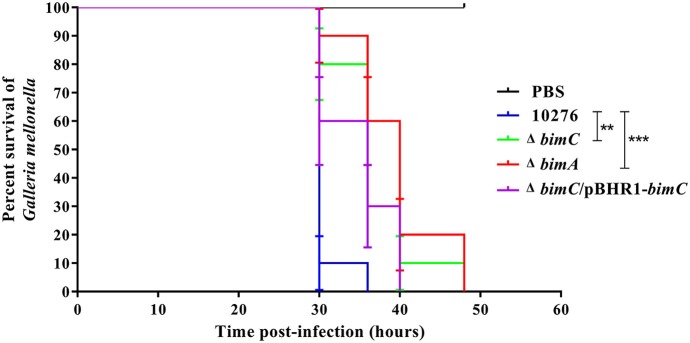
The intracellular pathogen Burkholderia pseudomallei, the etiological agent of melioidosis in humans and various animals, is capable of survival and movement within the cytoplasm of host cells by a process known as actin-based motility. The bacterial factor BimA is required for actin-based motility through its direct interaction with actin, and by mediating actin polymerization at a single pole of the bacterium to promote movement both within and between cells. However, little is known about the other bacterial proteins required for this process. Here, we have investigated the role of the bimC gene (bpss1491) which lies immediately upstream of the bimA gene (bpss1492) on the B. pseudomallei chromosome 2. Conserved amongst all B. pseudomallei, B. mallei and B. thailandensis strains sequenced to date, this gene encodes an iron-binding protein with homology to a group of proteins known as the bacterial autotransporter heptosyltransferase (BAHT) family. We have constructed a B. pseudomallei bimC deletion mutant and demonstrate that it is defective in intracellular survival in HeLa cells, but not in J774.1 macrophage-like cells. The bimC mutant is defective in cell to cell spread as demonstrated by ablation of plaque formation in HeLa cells, and by the inability to form multi-nucleated giant cells in J774.1 cells. These phenotypes in intracellular survival and cell to cell spread are not due to the loss of expression and polar localization of the BimA protein on the surface of intracellular bacteria, however they do correlate with an inability of the bacteria to recruit and polymerize actin. Furthermore, we also establish a role for bimC in virulence of B. pseudomallei using a Galleria mellonella larvae model of infection. Taken together, our findings indicate that B. pseudomallei BimC plays an important role in intracellular behavior and virulence of this emerging pathogen.
Keywords: BimA; BimC; Burkholderia pseudomallei; actin-based motility; intracellular survival; multi-nucleated giant cell; virulence.
Figures
References
-
- Boddey J. A., Day C. J., Flegg C. P., Ulrich R. L., Stephens S. R., Beacham I. R., et al. (2007). The bacteria‘l gene lfpA influences the potent induction of calcitonin receptor and osteoclast-related genes in Burkholderia pseudomallei-induced TRAP-positive multinucleated giant cells. Cell. Microbiol. 9, 514–531. 10.1111/j.1462-5822.2006.00807.x - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources