

Review
doi: 10.3389/fpls.2019.00360.
eCollection 2019.
Plant microRNAs: Biogenesis, Homeostasis, and Degradation
Affiliations
- PMID: 30972093
- PMCID: PMC6445950
- DOI: 10.3389/fpls.2019.00360
Item in Clipboard
Review
Plant microRNAs: Biogenesis, Homeostasis, and Degradation
Front Plant Sci.
.
Abstract
MicroRNAs (miRNAs), a class of endogenous, tiny, non-coding RNAs, are master regulators of gene expression among most eukaryotes. Intracellular miRNA abundance is regulated under multiple levels of control including transcription, processing, RNA modification, RNA-induced silencing complex (RISC) assembly, miRNA-target interaction, and turnover. In this review, we summarize our current understanding of the molecular components and mechanisms that influence miRNA biogenesis, homeostasis, and degradation in plants. We also make comparisons with findings from other organisms where necessary.
Keywords: Argonaute; DCL1; HEN1; miRNA biogenesis; target mimic; uridylation.
Figures
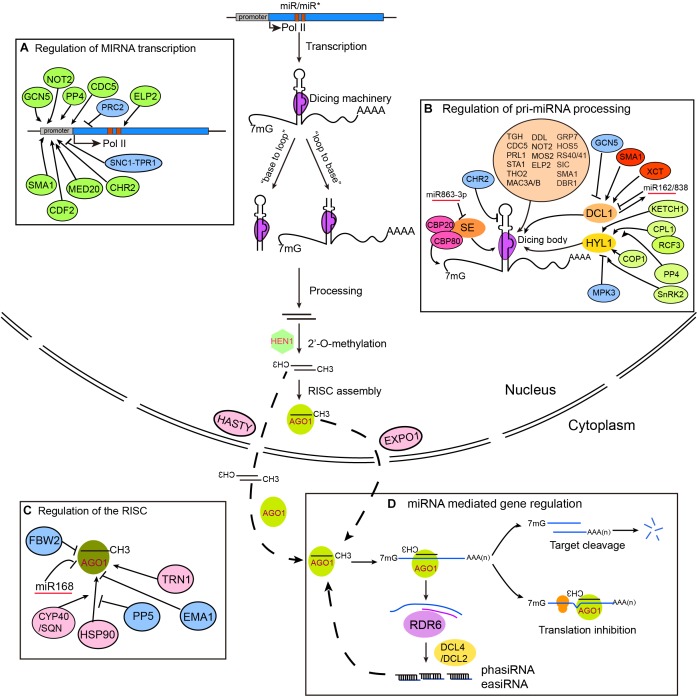
Regulation of miRNA biogenesis, RISC loading, and action in plants. MIRNA genes are transcribed by RNA Polymerase II (Pol II) and fold into stem-loop structures called pri-miRNAs. pri-miRNAs are mainly processed by DCL1 from either “base-to-loop” or “loop-to-base” direction. Nascent miRNA/miRNA∗ duplexes are methylated by the small RNA methyltransferase HEN1. RISC loading occurs in the nucleus, but may also occur in the cytoplasm. miRNAs mediate gene silencing through either target cleavage or translation inhibition. Some miRNAs can trigger the production of secondary siRNAs through sequential actions of RDR6 and DCL2/4, generating 21–22 nt secondary siRNAs called phasiRNAs and easiRNAs, which in turn repress gene expression via PTGS (for phasiRNAs) or TGS (for easiRNAs) (D). It is important to note that although the steps in the model are separate, they could be closely coupled. Factors involved in the regulation of MIR transcription, pri-miRNA processing, and RISC assembly are shown in parts (A–C), respectively.
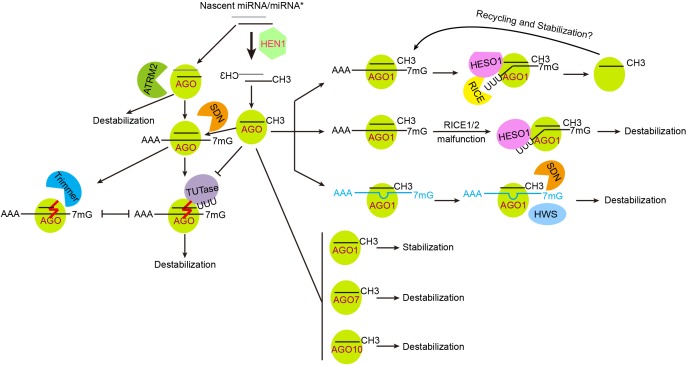
Regulation of miRNA stability and turnover. Plant miRNAs are heavily methylated, which is crucial for their stabilization. ATRM2 is involved in the degradation of unmethylated miRNA/miRNA∗s, likely during the initiation of RISC loading. RISC-associated unmethylated miRNAs are destabilized via 3′ end tailing and 3′-to-5′ trimming. TUTase, terminal uridylyl transferase such as HESO1 and URT1 in Arabidopsis and MUT68 in Chlamydomonas reinhardtii. Trimmer, a yet uncharacterized enzyme catalyzing 3′-to-5′ trimming of unmethylated miRNAs. The effects of AGO proteins on miRNA stability are judged only by the results of ago mutants and/or over-expression transgenic plants, and are not necessarily absolute. Defects in the degradation of cleavage products (e.g., in the RICE1/2 malfunction backgrounds) cause miRNA reduction, suggesting that successful release of cleavage products may facilitate RISC recycling and stabilization. TM (in blue color) induced miRNA destabilization involves the actions of SDNs and an F-box protein HWS.
References
-
- Achkar N. P., Cho S. K., Poulsen C., Arce A. L., Re D. A., Giudicatti A. J., et al. (2018). A Quick HYL1-dependent reactivation of MicroRNA production is required for a proper developmental response after extended periods of light deprivation. Dev Cell 46 236–247. 10.1016/j.devcel.2018.06.014 - DOI - PubMed
Publication types
LinkOut - more resources
Full Text Sources