

The phytochemical polydatin ameliorates non-alcoholic steatohepatitis by restoring lysosomal function and autophagic flux
- PMID: 30973211
- PMCID: PMC6533566
- DOI: 10.1111/jcmm.14320
The phytochemical polydatin ameliorates non-alcoholic steatohepatitis by restoring lysosomal function and autophagic flux
Abstract
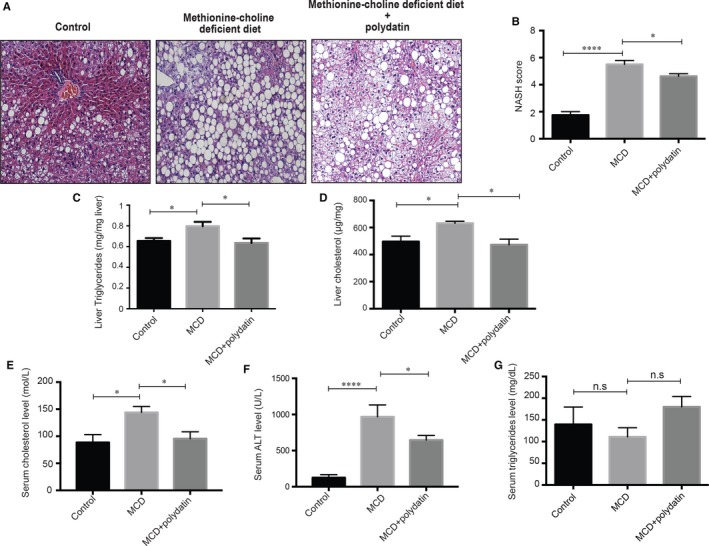
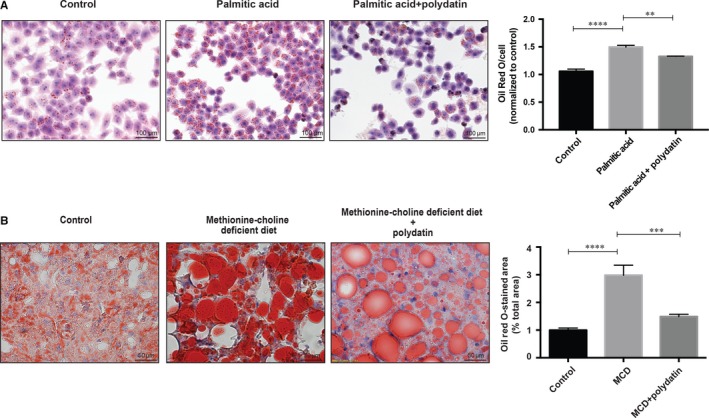
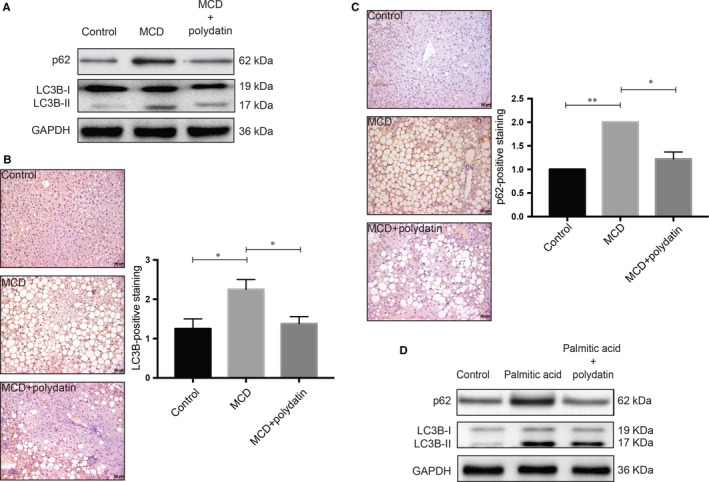
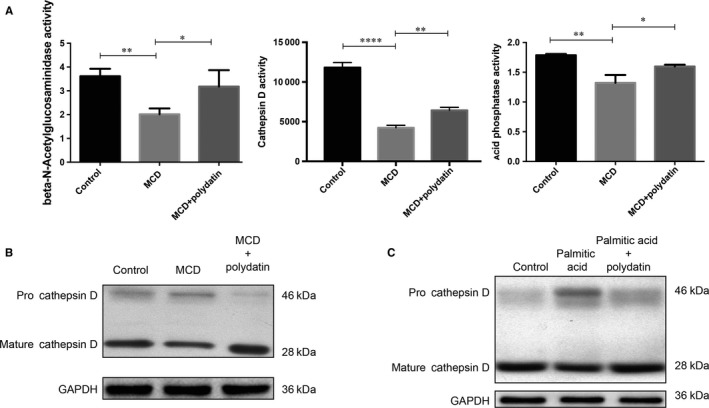
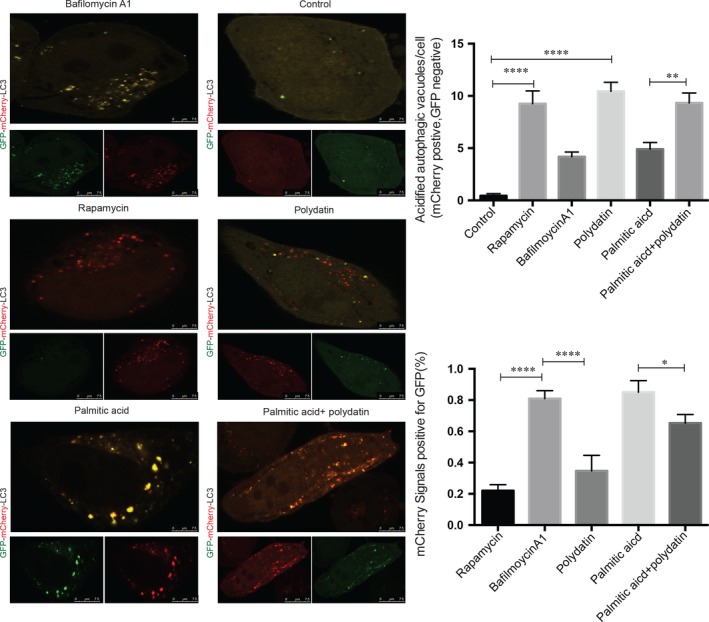
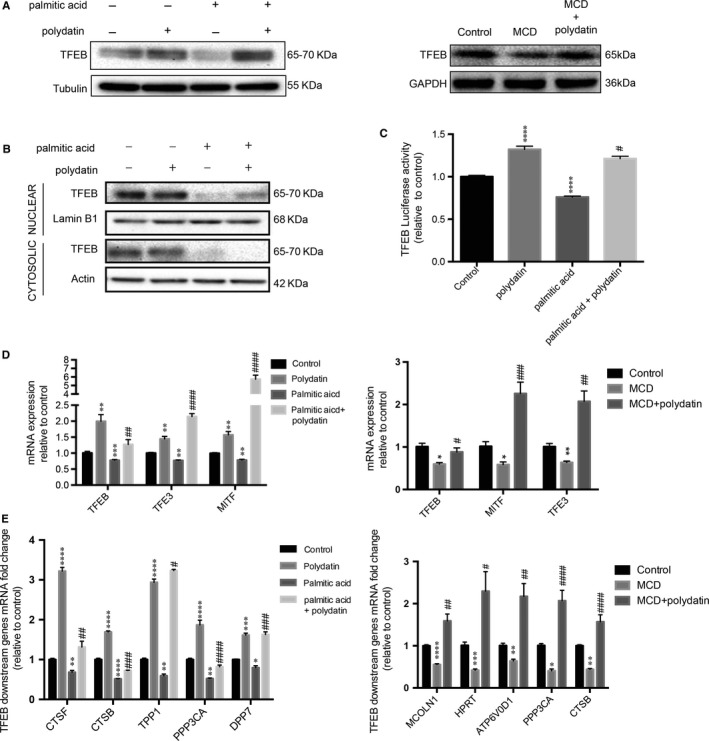
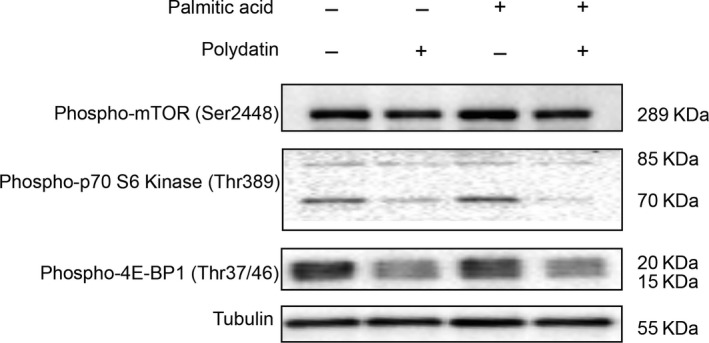
Impaired autophagic degradation of intracellular lipids is causally linked to the development of non-alcoholic steatohepatitis (NASH). Pharmacological agents that can restore hepatic autophagic flux could therefore have therapeutic potentials for this increasingly prevalent disease. Herein, we investigated the effects of polydatin, a natural precursor of resveratrol, in a murine nutritional model of NASH and a cell line model of steatosis. Results showed that oral administration of polydatin protected against hepatic lipid accumulation and alleviated inflammation and hepatocyte damage in db/db mice fed methionine-choline deficient diet. Polydatin also alleviated palmitic acid-induced lipid accumulation in cultured hepatocytes. In both models, polydatin restored lysosomal function and autophagic flux that were impaired by NASH or steatosis. Mechanistically, polydatin inhibited mTOR signalling and up-regulated the expression and activity of TFEB, a known master regulator of lysosomal function. In conclusion, polydatin ameliorated NASH through restoring autophagic flux. The polydatin-regulated autophagy was associated with inhibition of mTOR pathway and restoration of lysosomal function by TFEB. Our study provided affirmative preclinical evidence to inform future clinical trials for examining the potential anti-NASH effect of polydatin in humans.
Keywords: LC3; NAFLD; cathepsin D; lipophagy; p62.
© 2019 The Authors. Journal of Cellular and Molecular Medicine published by John Wiley & Sons Ltd and Foundation for Cellular and Molecular Medicine.
Conflict of interest statement
The authors confirm that there is no conflict of interest.
Figures
References
-
- Yeh MM, Brunt EM. Pathology of nonalcoholic fatty liver disease. Am J Clin Pathol. 2007;128(5):837‐847. - PubMed
-
- Yu J, Shen J, Sun TT, Zhang X, Wong N. Obesity, insulin resistance, NASH and hepatocellular carcinoma. Semin Cancer Biol 2013;23(6 Pt B):483‐491. - PubMed
-
- Neuschwander‐Tetri BA. Nontriglyceride hepatic lipotoxicity: the new paradigm for the pathogenesis of NASH. Curr Gastroenterol Rep. 2010;12(1):49‐56. - PubMed
-
- Wu W, Coffelt SB, Cho Ch, et al. The autophagic paradox in cancer therapy. Oncogene. 2012;31(8):939‐953. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
