

Contractile and hemodynamic forces coordinate Notch1b-mediated outflow tract valve formation
- PMID: 30973827
- PMCID: PMC6542601
- DOI: 10.1172/jci.insight.124460
Contractile and hemodynamic forces coordinate Notch1b-mediated outflow tract valve formation
Abstract
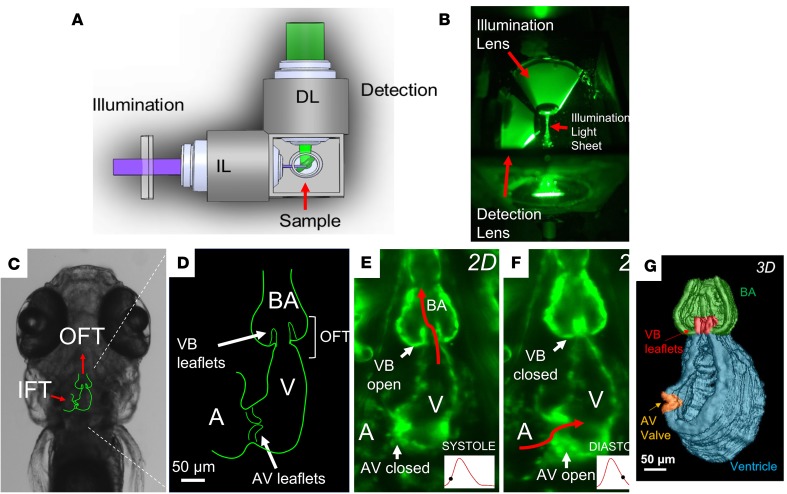
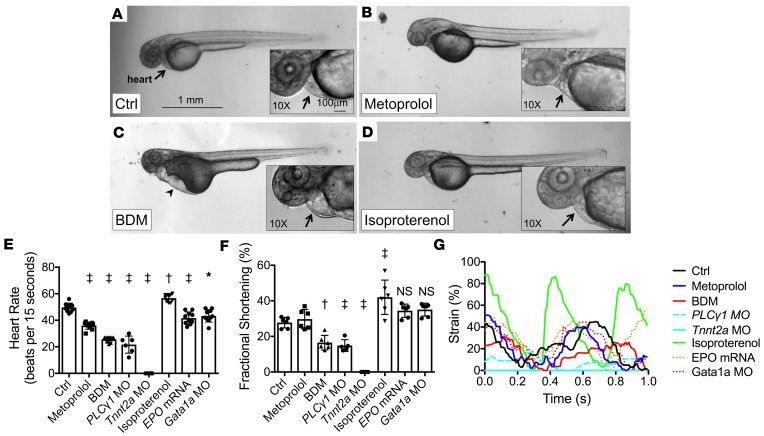
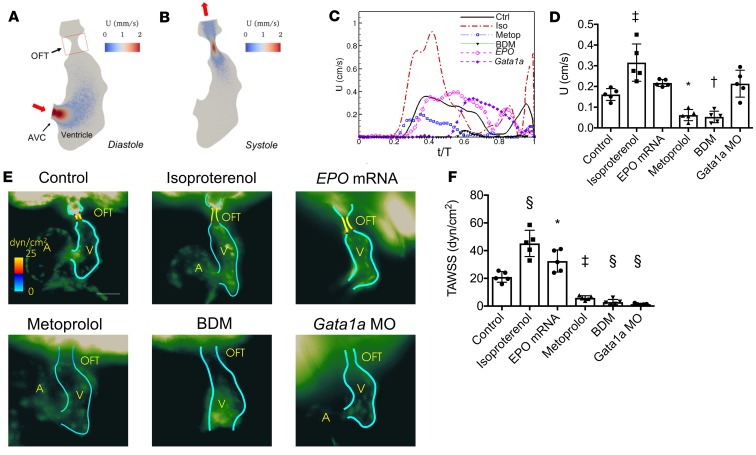
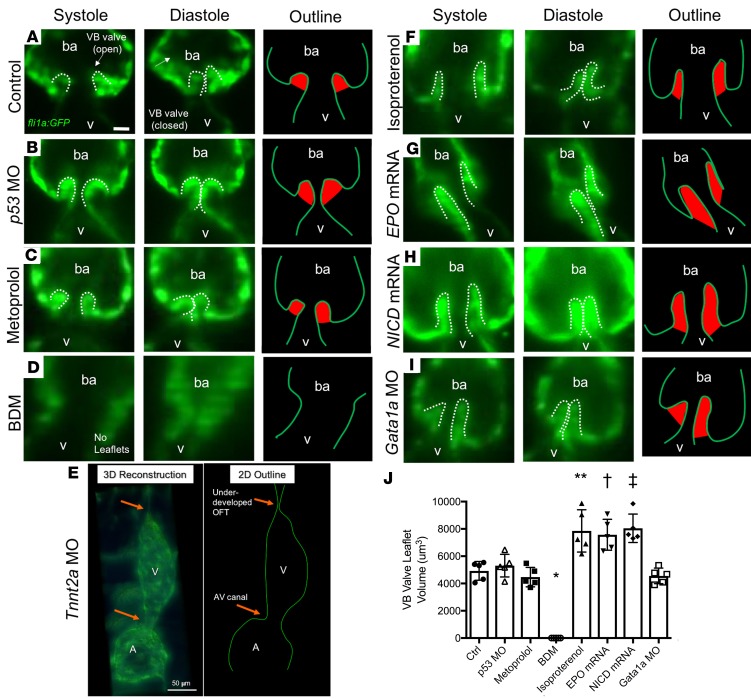
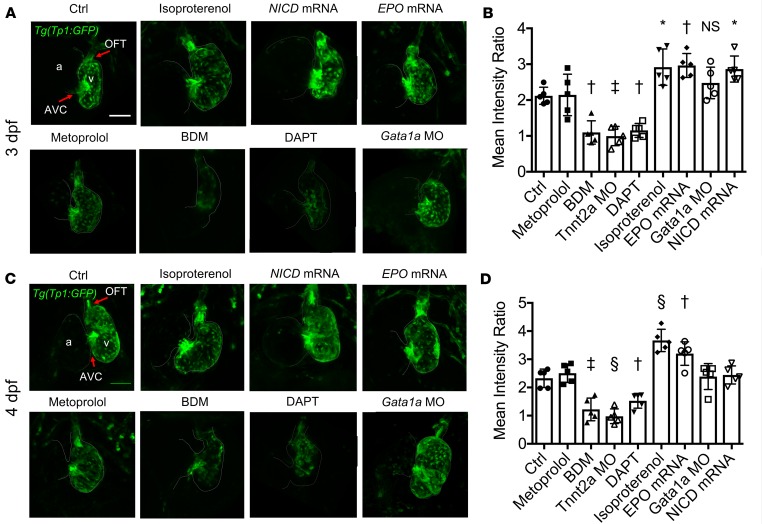
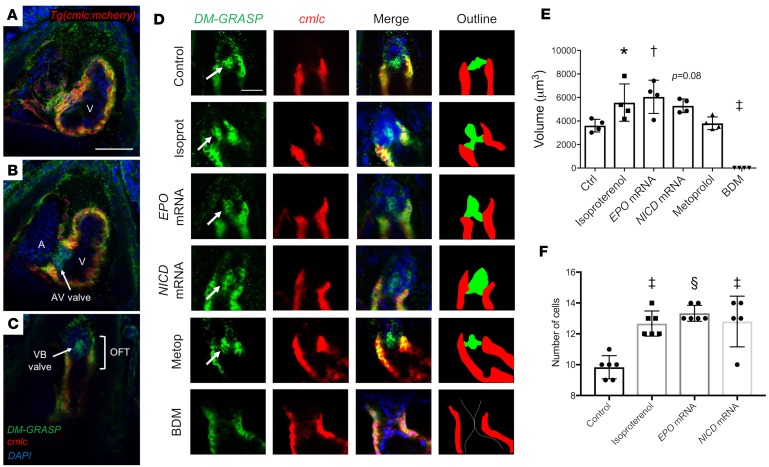
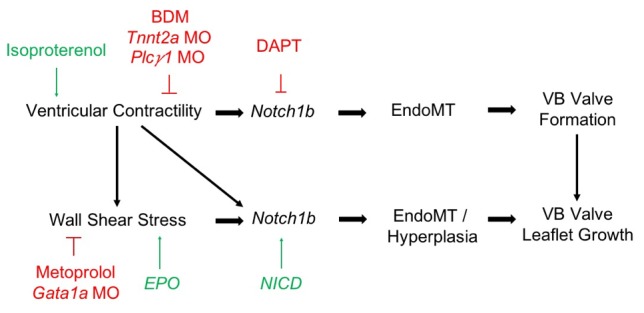
Biomechanical forces and endothelial-to-mesenchymal transition (EndoMT) are known to mediate valvulogenesis. However, the relative contributions of myocardial contractile and hemodynamic shear forces remain poorly understood. We integrated 4-D light-sheet imaging of transgenic zebrafish models with moving-domain computational fluid dynamics to determine effects of changes in contractile forces and fluid wall shear stress (WSS) on ventriculobulbar (VB) valve development. Augmentation of myocardial contractility with isoproterenol increased both WSS and Notch1b activity in the developing outflow tract (OFT) and resulted in VB valve hyperplasia. Increasing WSS in the OFT, achieved by increasing blood viscosity through EPO mRNA injection, also resulted in VB valve hyperplasia. Conversely, decreasing myocardial contractility by Tnnt2a morpholino oligonucleotide (MO) administration, 2,3-butanedione monoxime treatment, or Plcγ1 inhibition completely blocked VB valve formation, which could not be rescued by increasing WSS or activating Notch. Decreasing WSS in the OFT, achieved by slowing heart rate with metoprolol or reducing viscosity with Gata1a MO, did not affect VB valve formation. Immunofluorescent staining with the mesenchymal marker, DM-GRASP, revealed that biomechanical force-mediated Notch1b activity is implicated in EndoMT to modulate valve morphology. Altogether, increases in WSS result in Notch1b- EndoMT-mediated VB valve hyperplasia, whereas decreases in contractility result in reduced Notch1b activity, absence of EndoMT, and VB valve underdevelopment. Thus, we provide developmental mechanotransduction mechanisms underlying Notch1b-mediated EndoMT in the OFT.
Keywords: Cardiology; Cardiovascular disease; Development; Embryonic development.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
