

The early response of αB-crystallin to a single bout of aerobic exercise in mouse skeletal muscles depends upon fiber oxidative features
- PMID: 30974319
- PMCID: PMC6454247
- DOI: 10.1016/j.redox.2019.101183
The early response of αB-crystallin to a single bout of aerobic exercise in mouse skeletal muscles depends upon fiber oxidative features
Abstract
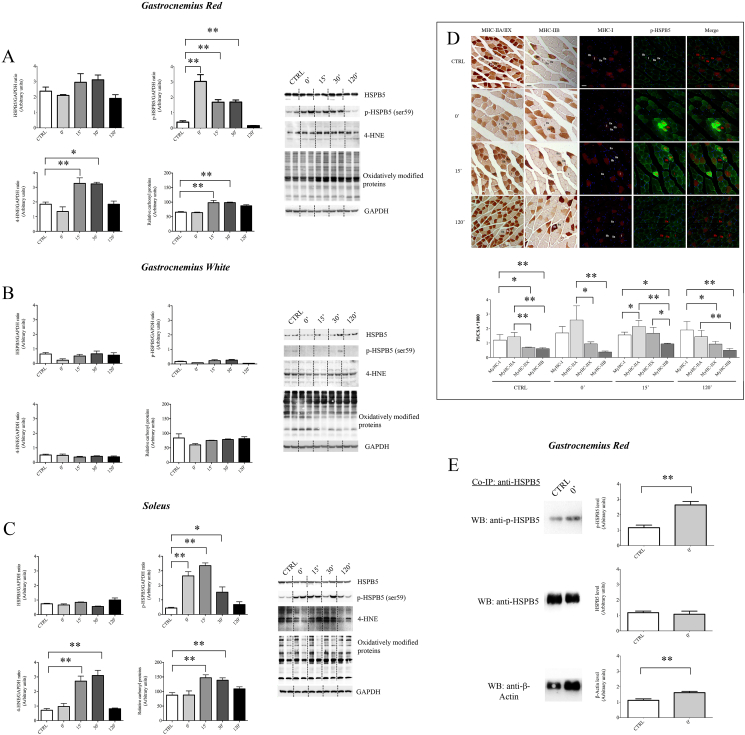
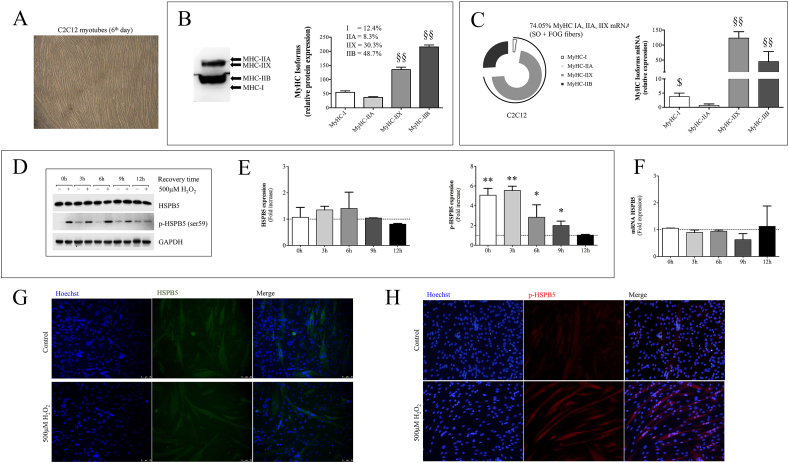
Besides its substantial role in eye lens, αB-crystallin (HSPB5) retains fundamental function in striated muscle during physiological or pathological modifications. In this study, we aimed to analyse the cellular and molecular factors driving the functional response of HSPB5 protein in different muscles from mice subjected to an acute bout of non-damaging endurance exercise or in C2C12 myocytes upon exposure to pro-oxidant environment, chosen as "in vivo" and "in vitro" models of a physiological stressing conditions, respectively. To this end, red (GR) and white gastrocnemius (GW), as sources of slow-oxidative and fast-glycolytic/oxidative fibers, as well as the soleus (SOL), mainly composed of slow-oxidative type fibers, were obtained from BALB/c mice, before (CTRL) and at different times (0', 15', 30' 120') following 1-h of running. Although the total level of HSPB5 protein was not affected by exercise, we found a significantly increase of phosphorylated HSPB5 (p-HSPB5) only in GR and SOL skeletal muscle with a higher amount of type I and IIA/X myofibers. The fiber-specific activation of HSPB5 was correlated to its interaction with the actin filaments, as well as to an increased level of lipid peroxidation and carbonylated proteins. The role of the pro-oxidant environment in HSPB5 response was investigated in terminally differentiated C2C12 myotubes, where most of HSPB5/pHSPB5 pool was present in the cytosolic compartment in standard culture conditions. As a result of exposure to pro-oxidizing, but not cytotoxic, H2O2 concentration, the p-38MAPK-mediated phosphorylation of HSPB5 resulted functional to promote its interaction with the myofibrillar components, such as β-actin, desmin and filamin 1. This study provides novel information on the molecular pathway underlying the HSPB5 physiological function in skeletal muscle, confirming the contribution of the pro-oxidant environment in HSPB5 activation and interaction with substrate/client myofibrillar proteins, offering new insights for the study of myofibrillar myopathies and cardiomyopathies.
Keywords: Endurance exercise; Oxidative stress; Skeletal muscle; αB-crystallin phosphorylation.
Copyright © 2019. Published by Elsevier B.V.
Figures







References
-
- Hartl F.U., Bracher A., Hayer-Hartl M. Molecular chaperones in protein folding and proteostasis. Nature. 2011;475:324–332. - PubMed
-
- Bakthisaran R., Tangirala R., Rao C.M. Small heat shock proteins: role in cellular functions and pathology. Biochim. Biophys. Acta. 2015;1854(4):291–319. - PubMed
-
- Benjamin I.J., Shelton J., Garry D.J., Richardson J.A. Temporospatial expression of the small HSP/alpha B-crystallin in cardiac and skeletal muscle during mouse development. Dev. Dynam. 1997;208:75–84. - PubMed
-
- Brady J.P., Garland D.L., Green D.E., Tamm E.R., Giblin F.J., Wawrousek E.F. AlphaB crystallin in lens development and muscle integrity: a gene knockout approach. Investig. Ophthalmol. Vis. Sci. 2001;42:2924–2934. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
