

Histone Methylation and Memory of Environmental Stress
- PMID: 30974922
- PMCID: PMC6523599
- DOI: 10.3390/cells8040339
Histone Methylation and Memory of Environmental Stress
Abstract
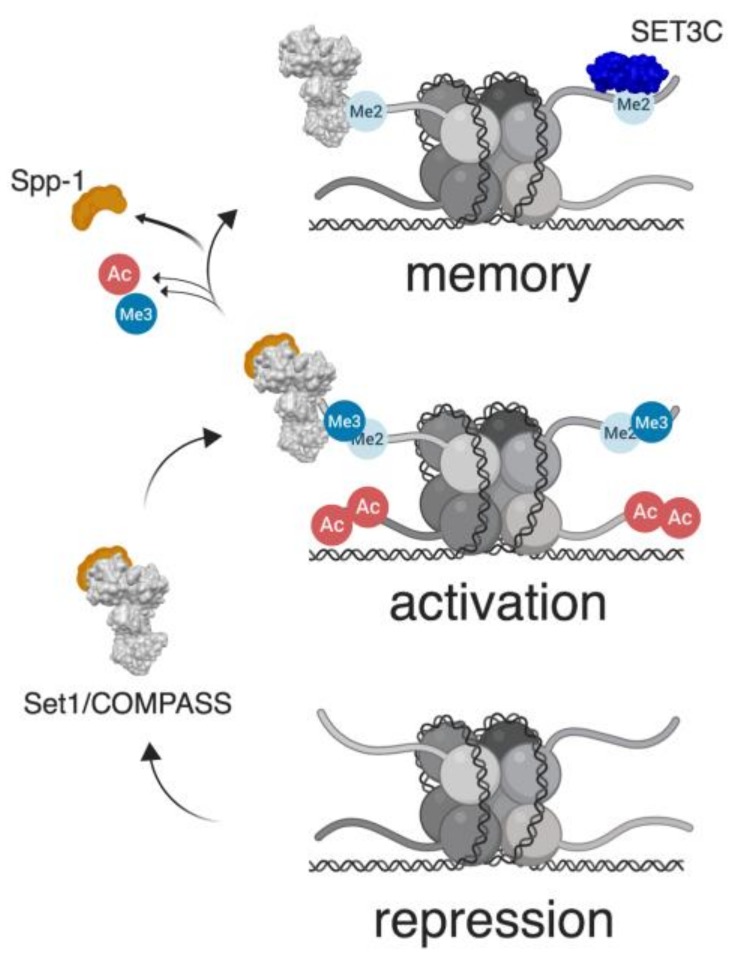
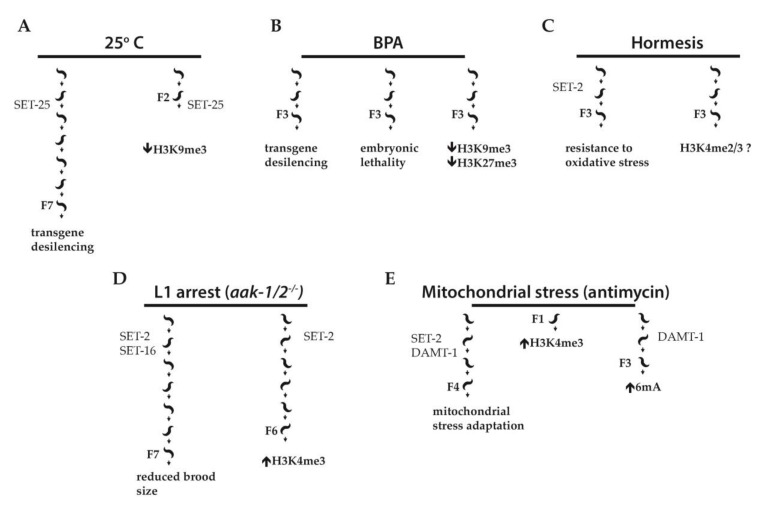
Cellular adaptation to environmental stress relies on a wide range of tightly controlled regulatory mechanisms, including transcription. Changes in chromatin structure and organization accompany the transcriptional response to stress, and in some cases, can impart memory of stress exposure to subsequent generations through mechanisms of epigenetic inheritance. In the budding yeast Saccharomyces cerevisiae, histone post-translational modifications, and in particular histone methylation, have been shown to confer transcriptional memory of exposure to environmental stress conditions through mitotic divisions. Recent evidence from Caenorhabditis elegans also implicates histone methylation in transgenerational inheritance of stress responses, suggesting a more widely conserved role in epigenetic memory.
Keywords: chromatin; epigenetic inheritance; histone methylation; stress; transcriptional memory.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Suganuma T., Workman J.L. Signals and combinatorial functions of histone modifications. Annu. Rev. Biochem. 2011;80:473–499. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
