

The three-dimensional structure of an H-superfamily conotoxin reveals a granulin fold arising from a common ICK cysteine framework
- PMID: 30975904
- PMCID: PMC6552430
- DOI: 10.1074/jbc.RA119.007491
The three-dimensional structure of an H-superfamily conotoxin reveals a granulin fold arising from a common ICK cysteine framework
Abstract
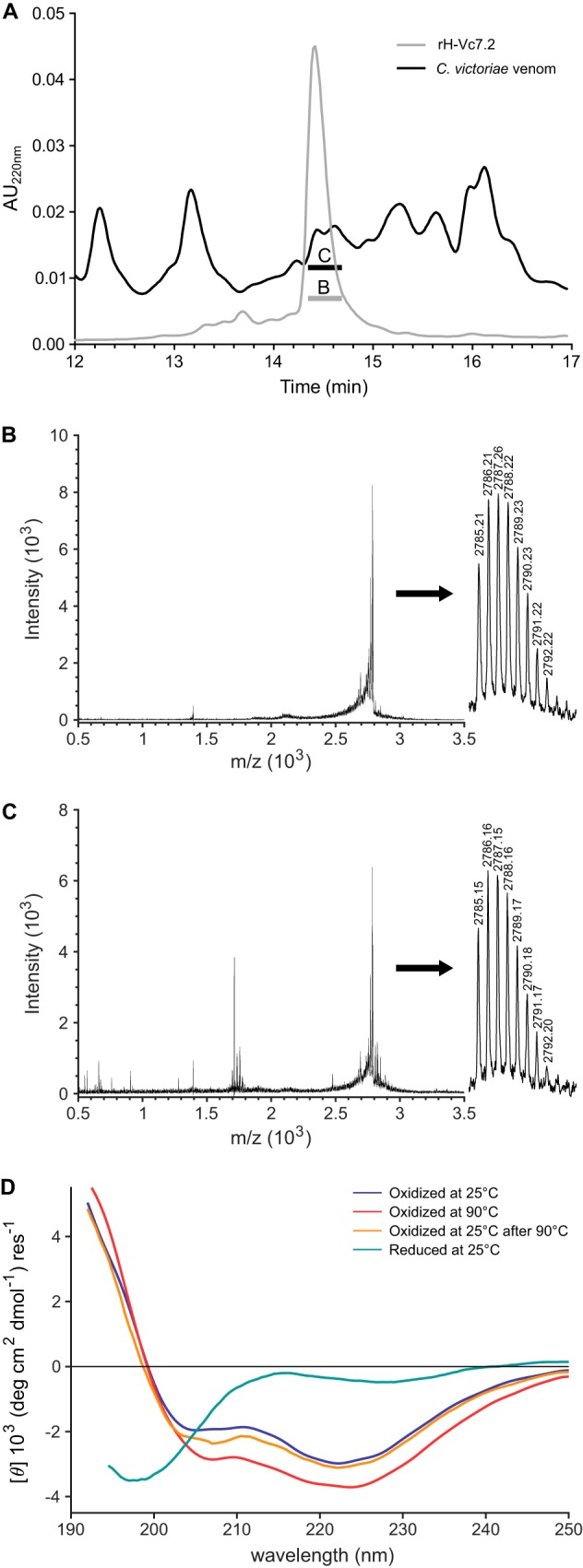
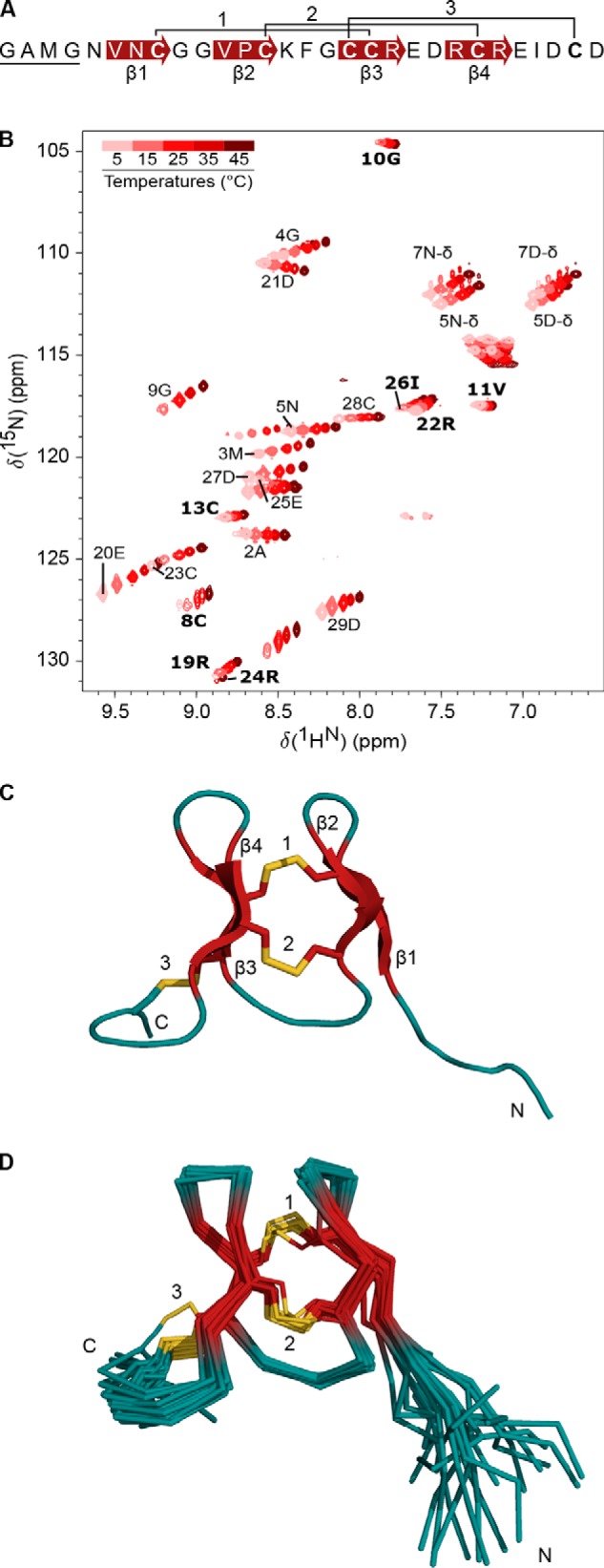
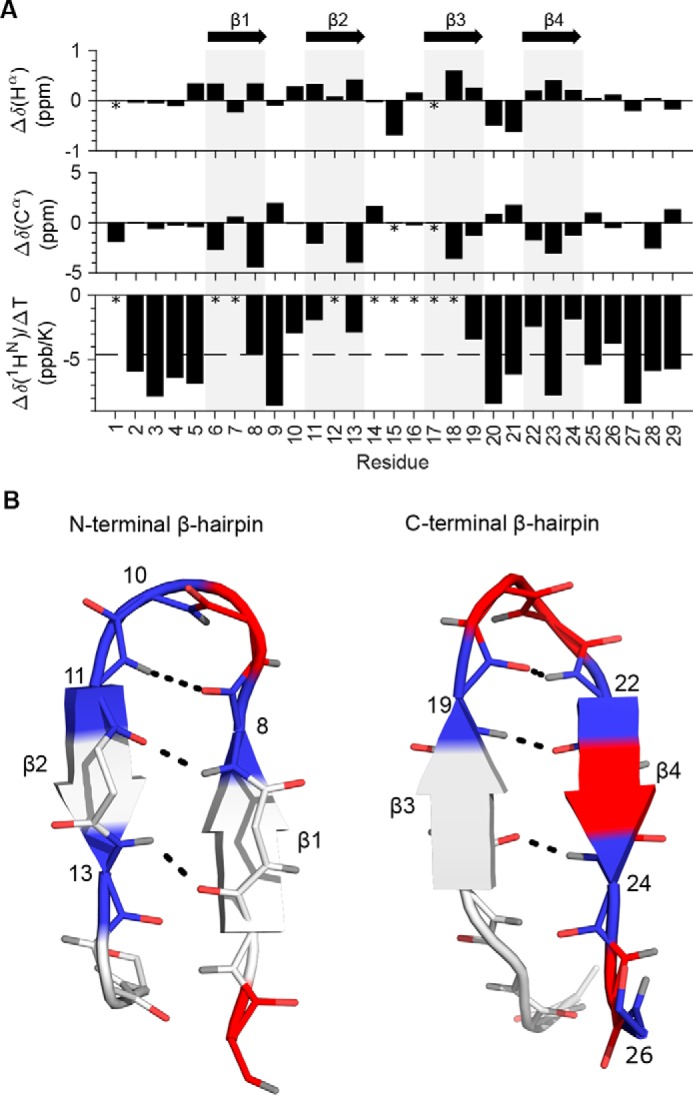
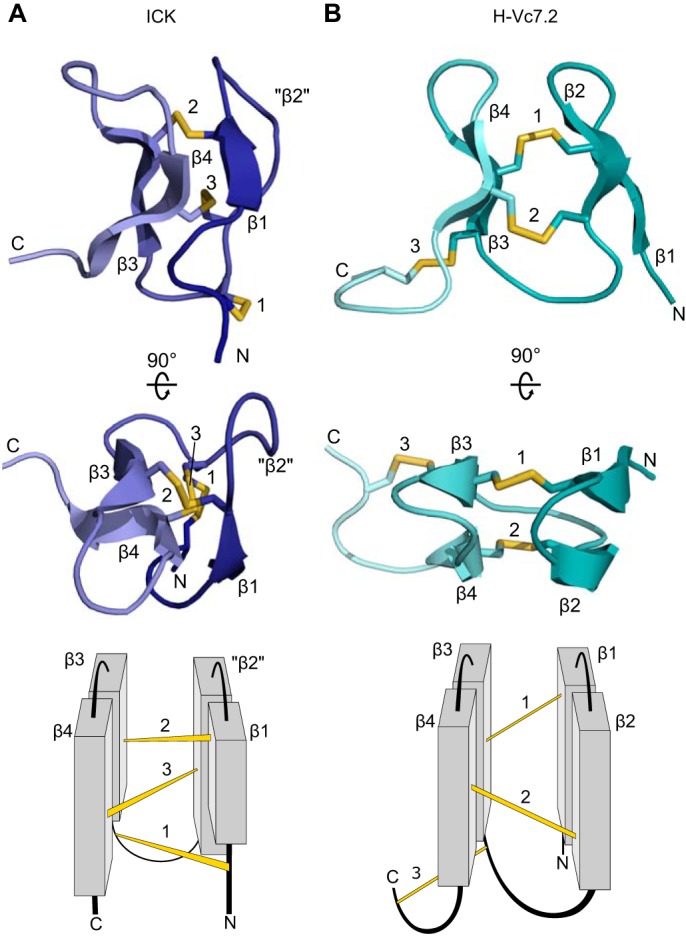
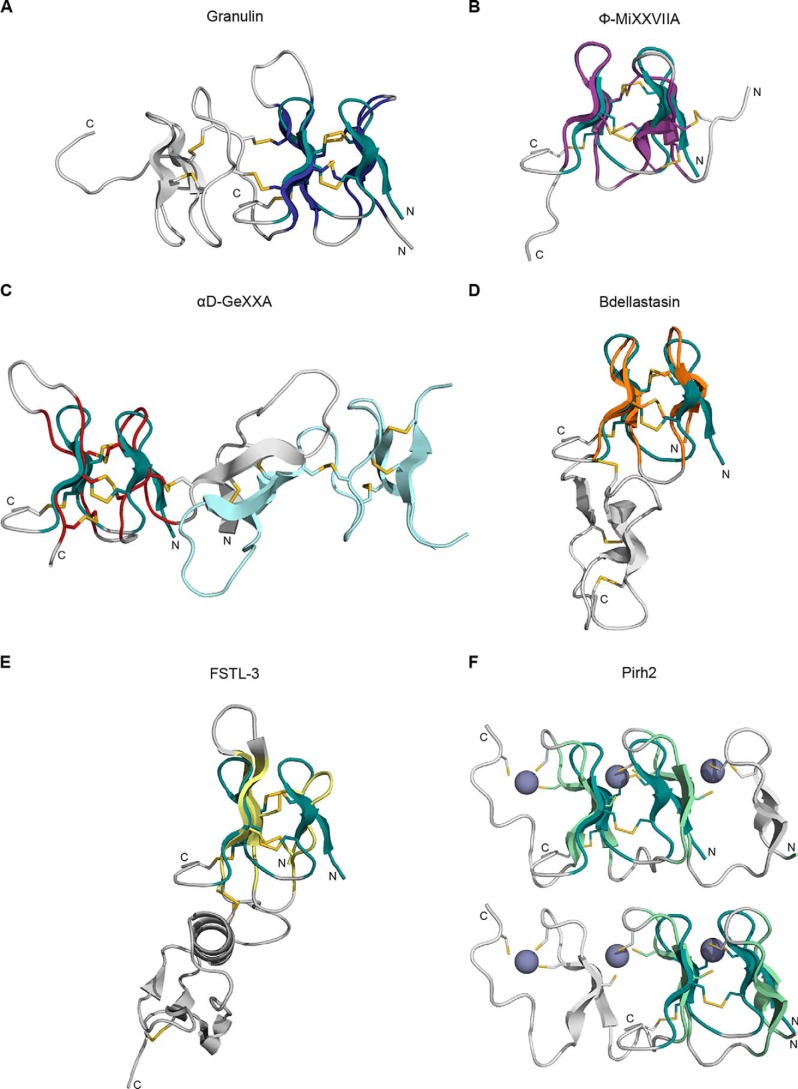
Venomous marine cone snails produce peptide toxins (conotoxins) that bind ion channels and receptors with high specificity and therefore are important pharmacological tools. Conotoxins contain conserved cysteine residues that form disulfide bonds that stabilize their structures. To gain structural insight into the large, yet poorly characterized conotoxin H-superfamily, we used NMR and CD spectroscopy along with MS-based analyses to investigate H-Vc7.2 from Conus victoriae, a peptide with a VI/VII cysteine framework. This framework has CysI-CysIV/CysII-CysV/CysIII-CysVI connectivities, which have invariably been associated with the inhibitor cystine knot (ICK) fold. However, the solution structure of recombinantly expressed and purified H-Vc7.2 revealed that although it displays the expected cysteine connectivities, H-Vc7.2 adopts a different fold consisting of two stacked β-hairpins with opposing β-strands connected by two parallel disulfide bonds, a structure homologous to the N-terminal region of the human granulin protein. Using structural comparisons, we subsequently identified several toxins and nontoxin proteins with this "mini-granulin" fold. These findings raise fundamental questions concerning sequence-structure relationships within peptides and proteins and the key determinants that specify a given fold.
Keywords: CD spectroscopy; NMR spectroscopy; antistasin; conotoxin; disulfide; disulfide bond; granulin; inhibitor cystine knot; protein conformation; protein evolution; protein expression; protein structure; protein-disulfide isomerase; toxin; β-hairpin.
© 2019 Nielsen et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
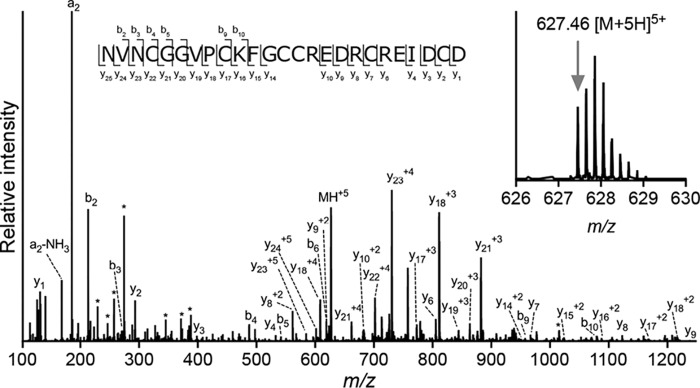
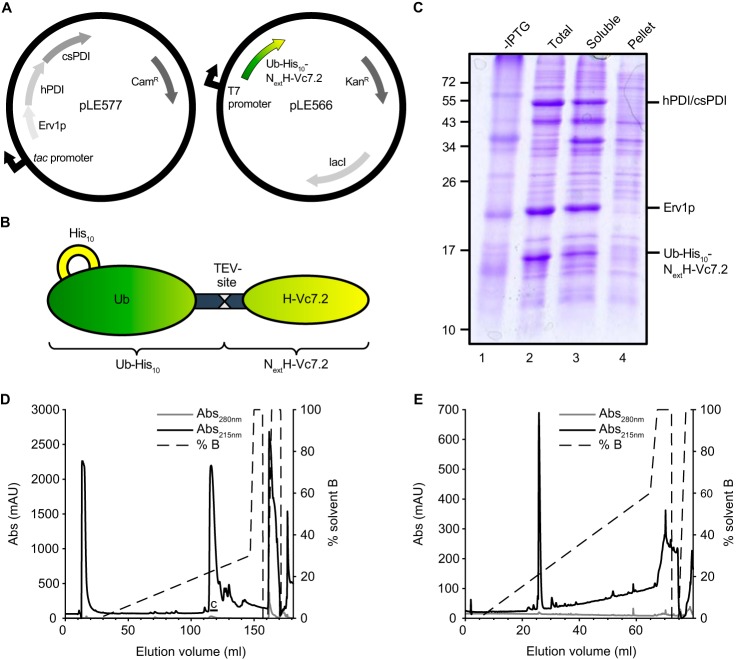
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
