

Cell-intrinsic depletion of Aml1-ETO-expressing pre-leukemic hematopoietic stem cells by K-Ras activating mutation
- PMID: 30975913
- PMCID: PMC6821613
- DOI: 10.3324/haematol.2018.205351
Cell-intrinsic depletion of Aml1-ETO-expressing pre-leukemic hematopoietic stem cells by K-Ras activating mutation
Abstract
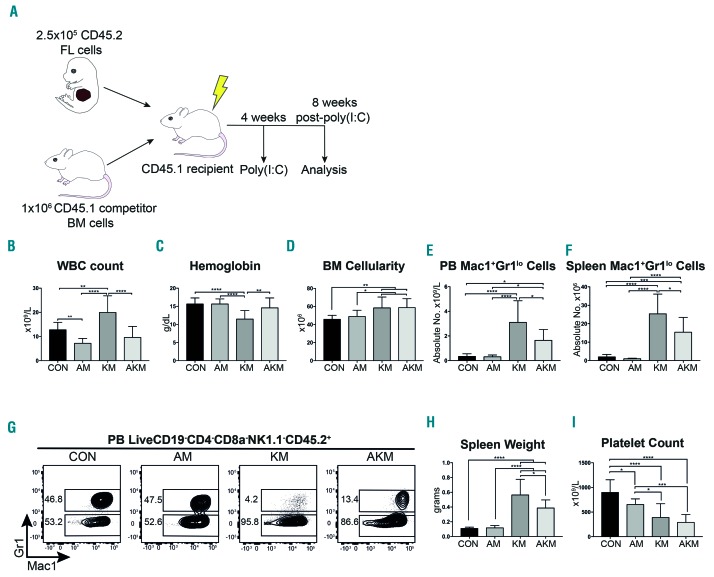
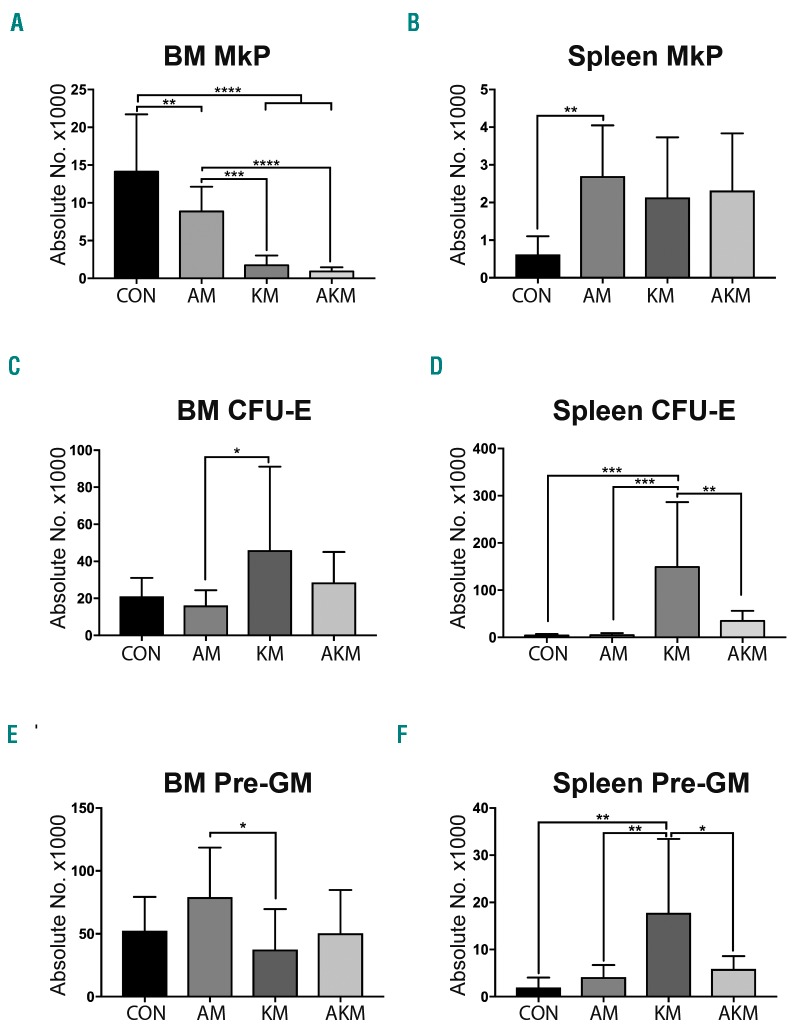
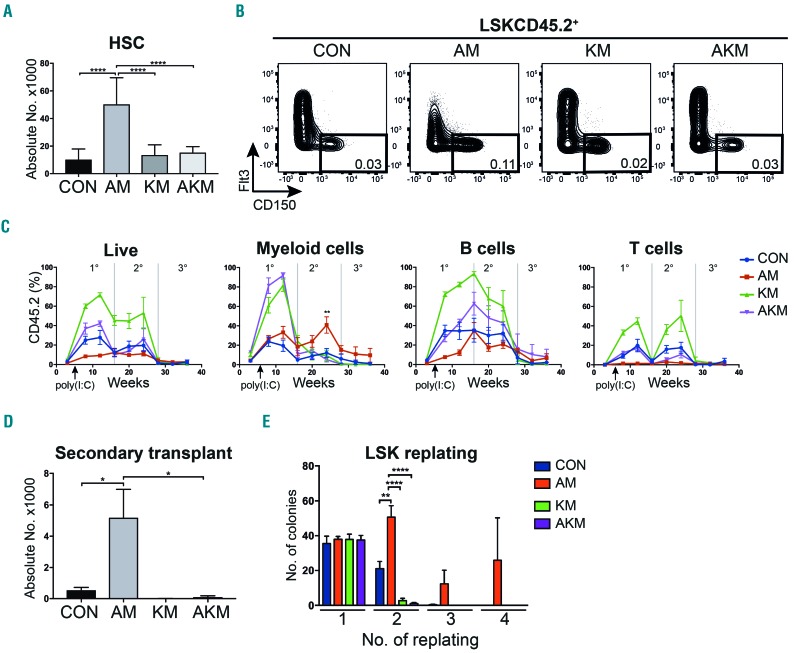
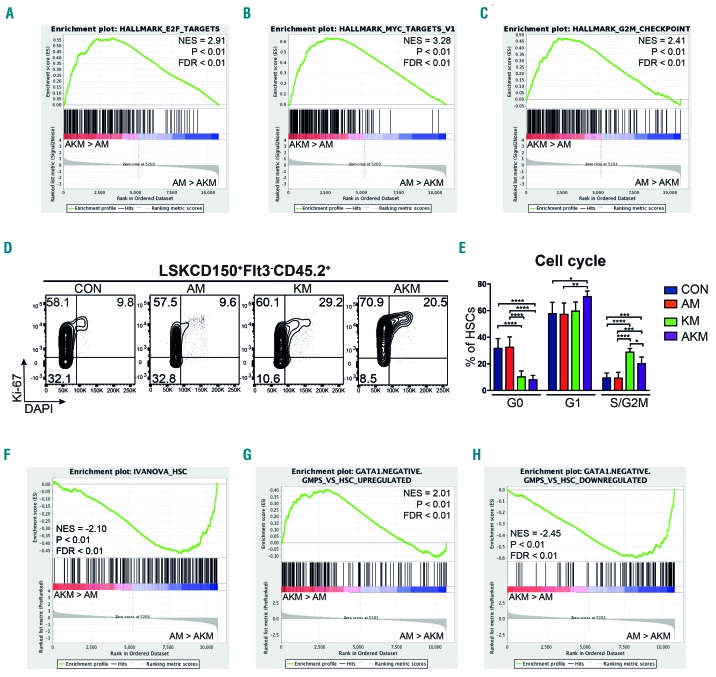
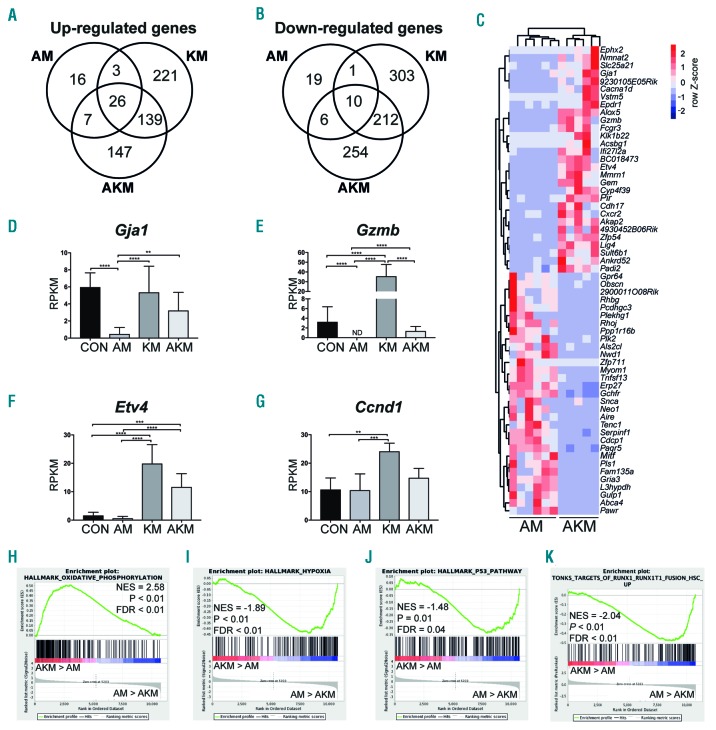
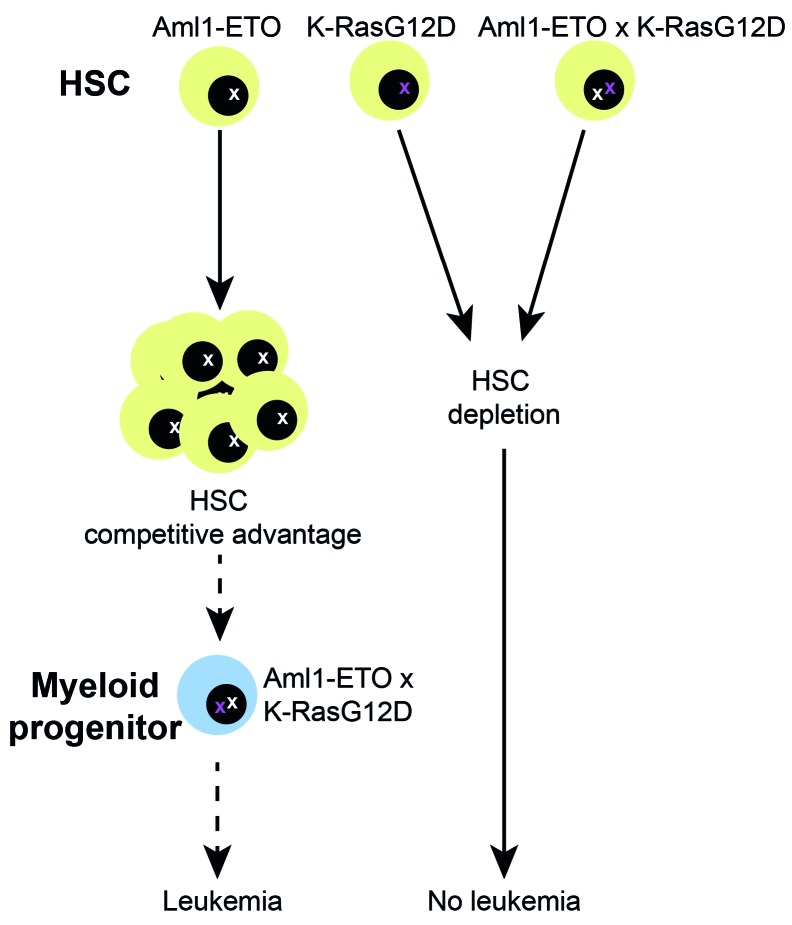
Somatic mutations in acute myeloid leukemia are acquired sequentially and hierarchically. First, pre-leukemic mutations, such as t(8;21) that encodes AML1-ETO, are acquired within the hematopoietic stem cell (HSC) compartment, while signaling pathway mutations, including KRAS activating mutations, are late events acquired during transformation of leukemic progenitor cells and are rarely detectable in HSC. This raises the possibility that signaling pathway mutations are detrimental to clonal expansion of pre-leukemic HSC. To address this hypothesis, we used conditional genetics to introduce Aml1-ETO and K-RasG12D into murine HSC, either individually or in combination. In the absence of activated Ras, Aml1-ETO-expressing HSC conferred a competitive advantage. However, activated K-Ras had a marked detrimental effect on Aml1-ETO-expressing HSC, leading to loss of both phenotypic and functional HSC. Cell cycle analysis revealed a loss of quiescence in HSC co-expressing Aml1-ETO and K-RasG12D, accompanied by an enrichment in E2F and Myc target gene expression and depletion of HSC self-renewal-associated gene expression. These findings provide a mechanistic basis for the observed absence of KRAS signaling mutations in the pre-malignant HSC compartment.
Copyright© 2019 Ferrata Storti Foundation.
Figures
Comment in
-
Standing at odds: mutated RAS and hematopoietic stem cells.Haematologica. 2019 Nov;104(11):2125-2128. doi: 10.3324/haematol.2019.230029. Haematologica. 2019. PMID: 31666341 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
