

AIRE expression controls the peripheral selection of autoreactive B cells
- PMID: 30979797
- PMCID: PMC7257641
- DOI: 10.1126/sciimmunol.aav6778
AIRE expression controls the peripheral selection of autoreactive B cells
Abstract
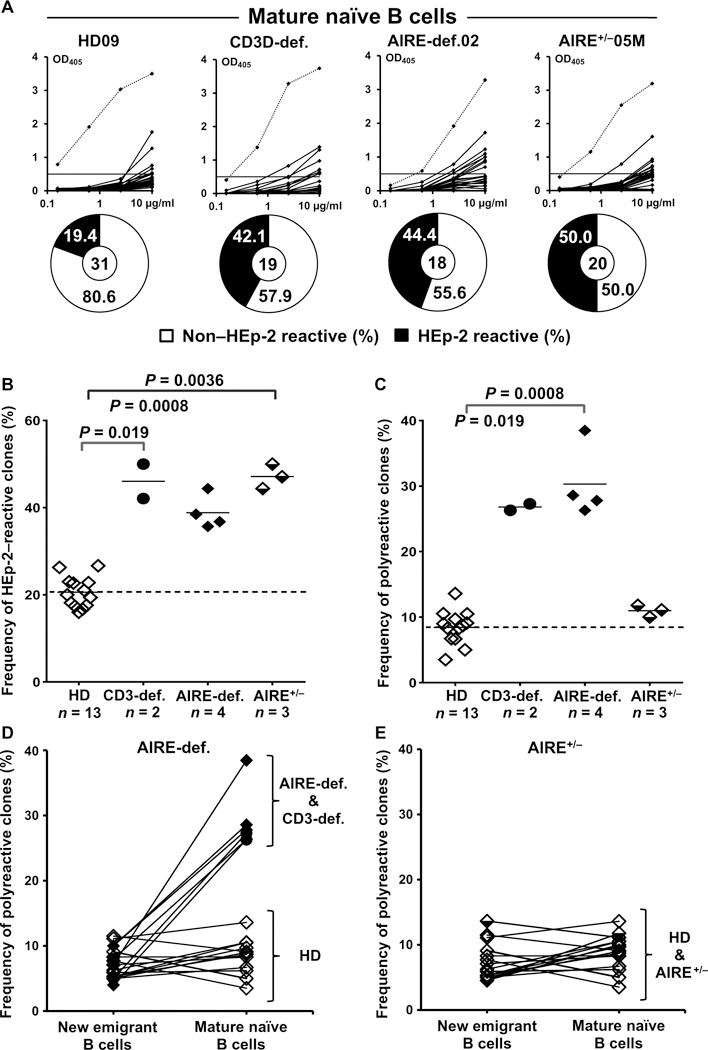
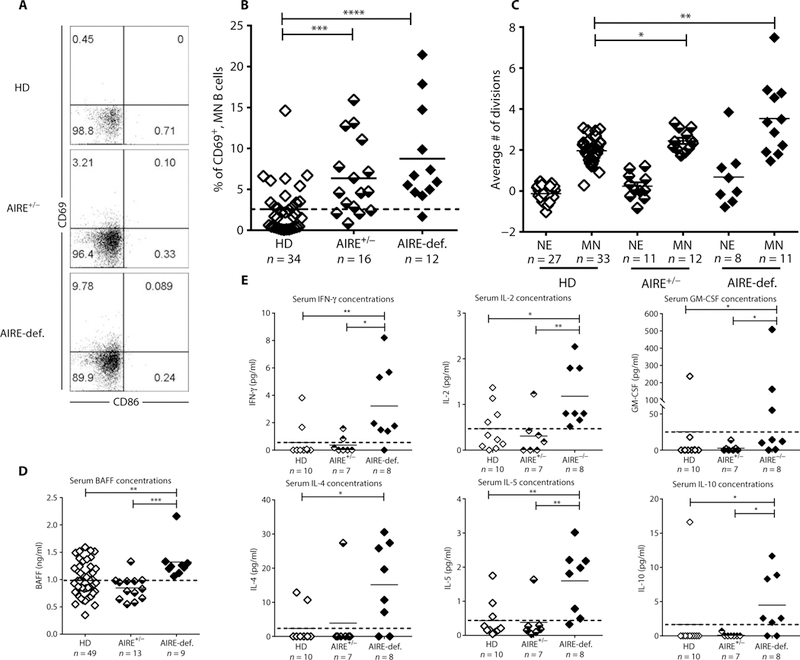
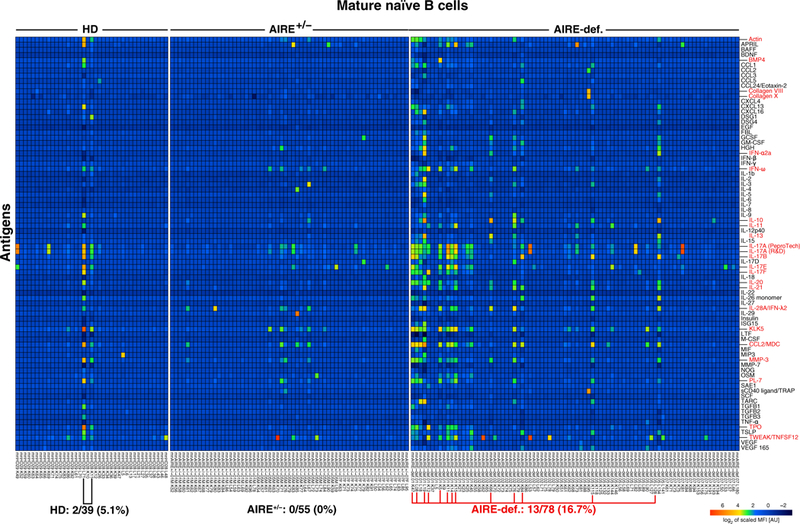
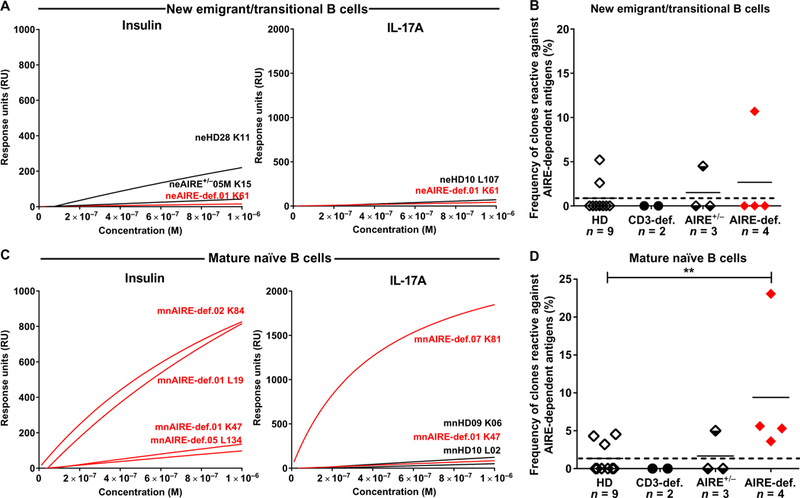
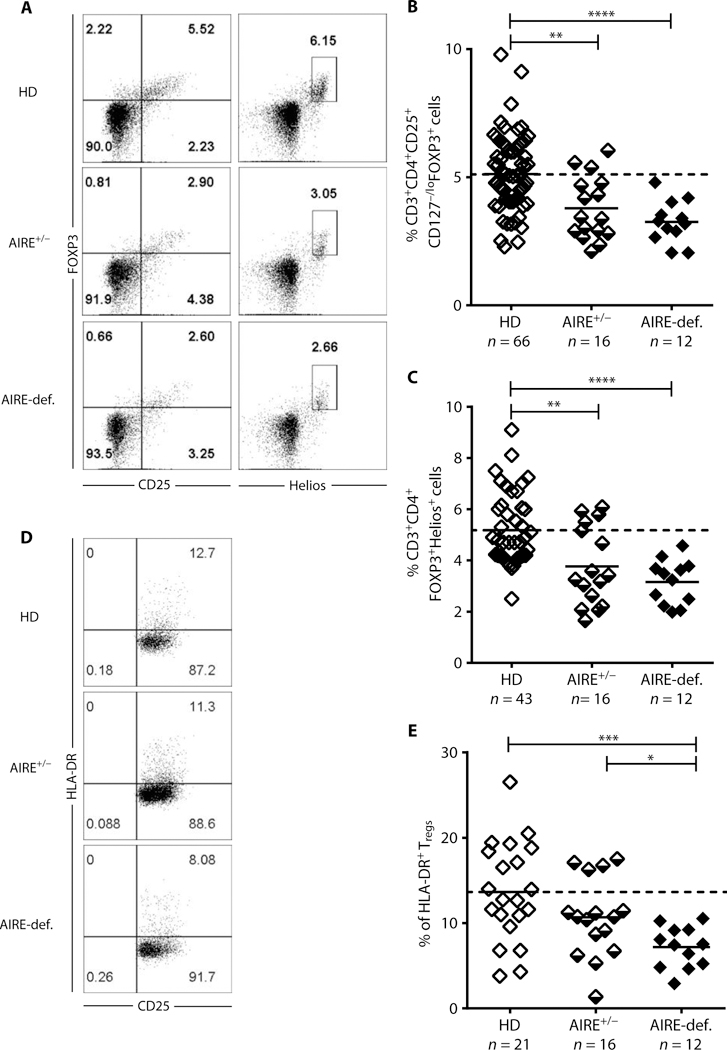
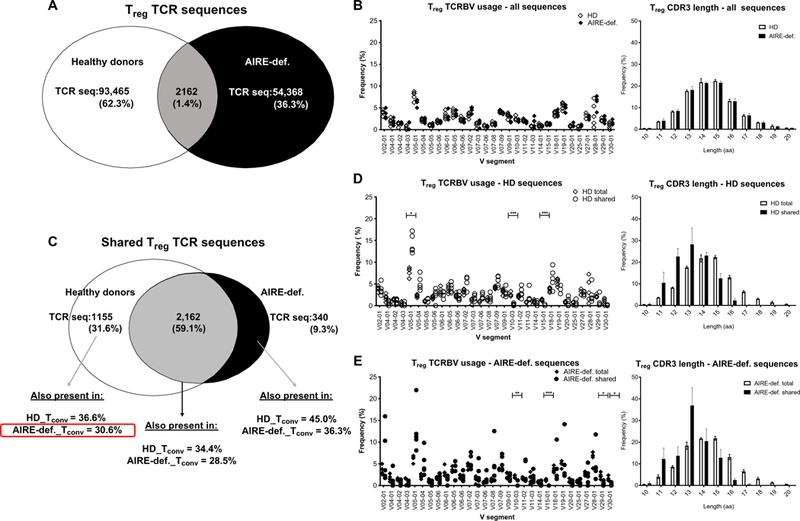
Autoimmune regulator (AIRE) mutations result in autoimmune polyendocrinopathy-candidiasis-ectodermal dystrophy (APECED) syndrome characterized by defective central T cell tolerance and the production of many autoantibodies targeting tissue-specific antigens and cytokines. By studying CD3- and AIRE-deficient patients, we found that lack of either T cells or AIRE function resulted in the peripheral accumulation of autoreactive mature naïve B cells. Proteomic arrays and Biacore affinity measurements revealed that unmutated antibodies expressed by these autoreactive naïve B cells recognized soluble molecules and cytokines including insulin, IL-17A, and IL-17F, which are AIRE-dependent thymic peripheral tissue antigens targeted by autoimmune responses in APECED. AIRE-deficient patients also displayed decreased frequencies of regulatory T cells (Tregs) that lacked common TCRβ clones found instead in their conventional T cell compartment, thereby suggesting holes in the Treg TCR repertoire of these patients. Hence, AIRE-mediated T cell/Treg selection normally prevents the expansion of autoreactive naïve B cells recognizing peripheral self-antigens.
Copyright © 2019 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Figures

References
-
- Nagamine K, Peterson P, Scott HS, Kudoh J, Minoshima S, Heino M, Krohn KJE, Lalioti MD, Mullis PE, Antonarakis SE, Kawasaki K, Asakawa S, Ito F, Shimizu N, Positional cloning of the APECED gene. Nat. Genet. 17, 393–398 (1997). - PubMed
-
- Finnish-German APECED Consortium, An autoimmune disease, APECED, caused by mutations in a novel gene featuring two PHD-type zinc-finger domains. Nat. Genet. 17, 399–403 (1997). - PubMed
-
- Anderson MS, Venanzi ES, Klein L, Chen Z, Berzins SP, Turley SJ, von Boehmer H, Bronson R, Dierich A, Benoist C, Mathis D, Projection of an immunological self shadow within the thymus by the aire protein. Science 298, 1395–1401 (2002). - PubMed
-
- Kärner J, Meager A, Laan M, Maslovskaja J, Pihlap M, Remm A, Juronen E, Wolff ASB, Husebye ES, Podkrajsek KT, Bratanic N, Battelino T, Willcox N, Peterson P, Kisand K, Anti-cytokine autoantibodies suggest pathogenetic links with autoimmune regulator deficiency in humans and mice. Clin. Exp. Immunol. 171, 263–272 (2013). - PMC - PubMed
-
- Ramsey C, Winqvist O, Puhakka L, Halonen M, Moro A, Kämpe O, Eskelin P, Pelto-Huikko M, Peltonen L, Aire deficient mice develop multiple features of APECED phenotype and show altered immune response. Hum. Mol. Genet. 11, 397–409 (2002). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
