

Deep-Subsurface Pressure Stimulates Metabolic Plasticity in Shale-Colonizing Halanaerobium spp
- PMID: 30979840
- PMCID: PMC6544827
- DOI: 10.1128/AEM.00018-19
Deep-Subsurface Pressure Stimulates Metabolic Plasticity in Shale-Colonizing Halanaerobium spp
Abstract
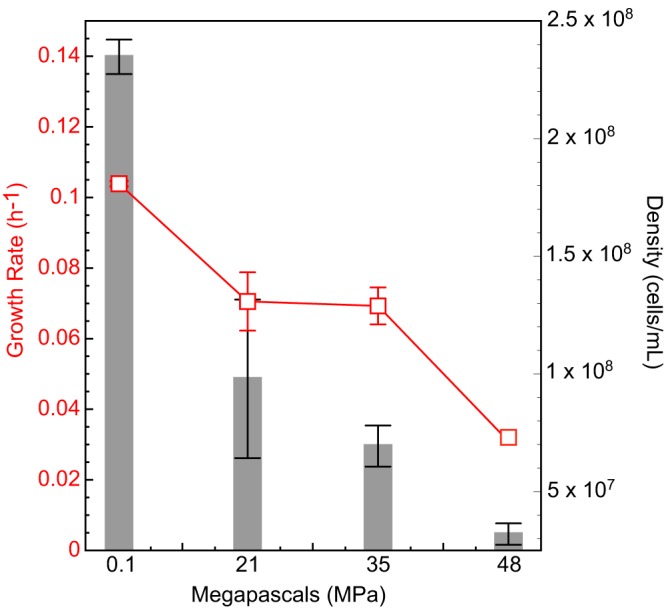
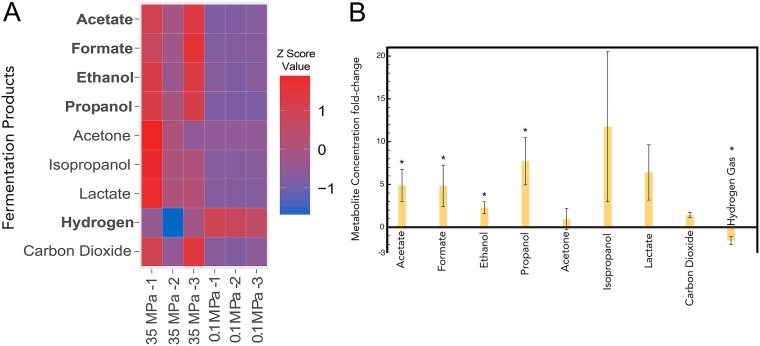
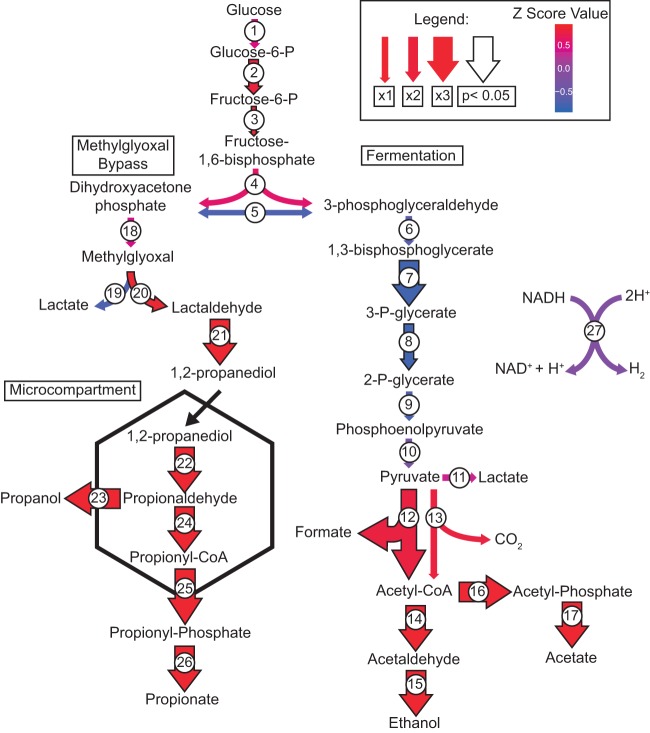
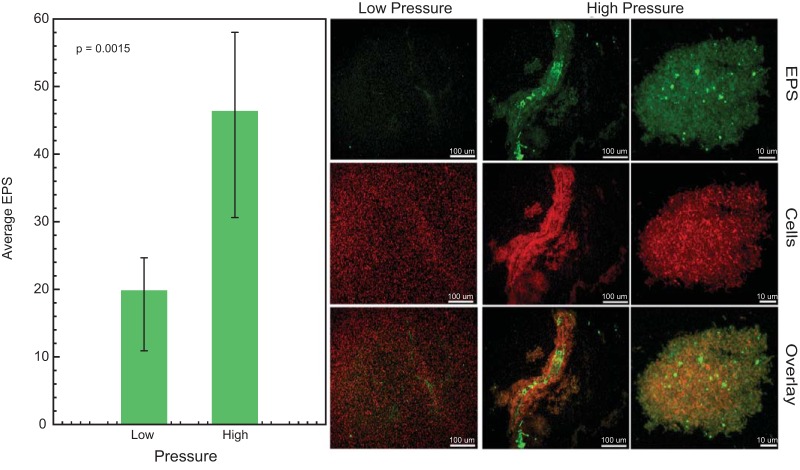
Bacterial Halanaerobium strains become the dominant persisting microbial community member in produced fluids across geographically distinct hydraulically fractured shales. Halanaerobium is believed to be inadvertently introduced into this environment during the drilling and fracturing process and must therefore tolerate large changes in pressure, temperature, and salinity. Here, we used a Halanaerobium strain isolated from a natural gas well in the Utica Point Pleasant formation to investigate metabolic and physiological responses to growth under high-pressure subsurface conditions. Laboratory incubations confirmed the ability of Halanaerobium congolense strain WG8 to grow under pressures representative of deep shale formations (21 to 48 MPa). Under these conditions, broad metabolic and physiological shifts were identified, including higher abundances of proteins associated with the production of extracellular polymeric substances. Confocal laser scanning microscopy indicated that extracellular polymeric substance (EPS) production was associated with greater cell aggregation when biomass was cultured at high pressure. Changes in Halanaerobium central carbon metabolism under the same conditions were inferred from nuclear magnetic resonance (NMR) and gas chromatography measurements, revealing large per-cell increases in production of ethanol, acetate, and propanol and cessation of hydrogen production. These metabolic shifts were associated with carbon flux through 1,2-propanediol in response to slower fluxes of carbon through stage 3 of glycolysis. Together, these results reveal the potential for bioclogging and corrosion (via organic acid fermentation products) associated with persistent Halanaerobium growth in deep, hydraulically fractured shale ecosystems, and offer new insights into cellular mechanisms that enable these strains to dominate deep-shale microbiomes.IMPORTANCE The hydraulic fracturing of deep-shale formations for hydrocarbon recovery accounts for approximately 60% of U.S. natural gas production. Microbial activity associated with this process is generally considered deleterious due to issues associated with sulfide production, microbially induced corrosion, and bioclogging in the subsurface. Here we demonstrate that a representative Halanaerobium species, frequently the dominant microbial taxon in hydraulically fractured shales, responds to pressures characteristic of the deep subsurface by shifting its metabolism to generate more corrosive organic acids and produce more polymeric substances that cause "clumping" of biomass. While the potential for increased corrosion of steel infrastructure and clogging of pores and fractures in the subsurface may significantly impact hydrocarbon recovery, these data also offer new insights for microbial control in these ecosystems.
Keywords: Halanaerobium; biofilms; high pressure; hydraulic fracturing; metabolomics; shale.
Copyright © 2019 Booker et al.
Figures


References
-
- Daly RA, Borton MA, Wilkins MJ, Hoyt DW, Kountz DJ, Wolfe RA, Welch SA, Marcus DN, Trexler RV, MacRae JD, Krzycki JA, Cole DR, Mouser PJ, Wrighton KC. 2016. Microbial metabolisms in a 2.5-km-deep ecosystem created by hydraulic fracturing in shales. Nat Microbiol 1:16146. doi: 10.1038/nmicrobiol.2016.146. - DOI - PubMed
-
- Booker AE, Borton MA, Daly RA, Welch SA, Nicora CD, Hoyt DW, Wilson T, Purvine SO, Wolfe RA, Sharma S, Mouser PJ, Cole DR, Lipton MS, Wrighton KC, Wilkins MJ. 2017. Sulfide generation by dominant Halanaerobium microorganisms in hydraulically fractured shales. mSphere 2:e00257-17. doi: 10.1128/mSphereDirect.00257-17. - DOI - PMC - PubMed
Publication types
MeSH terms
Supplementary concepts
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
