

Signatures of Divergence, Invasiveness, and Terrestrialization Revealed by Four Apple Snail Genomes
- PMID: 30980073
- PMCID: PMC6573481
- DOI: 10.1093/molbev/msz084
Signatures of Divergence, Invasiveness, and Terrestrialization Revealed by Four Apple Snail Genomes
Abstract
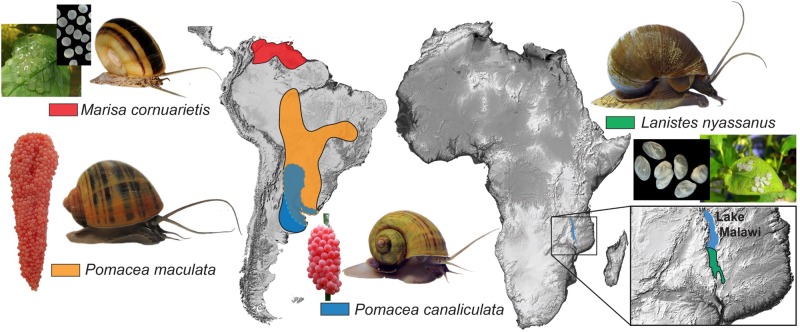
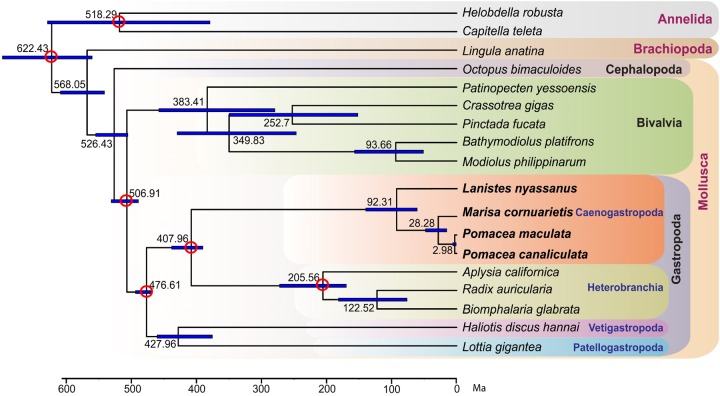
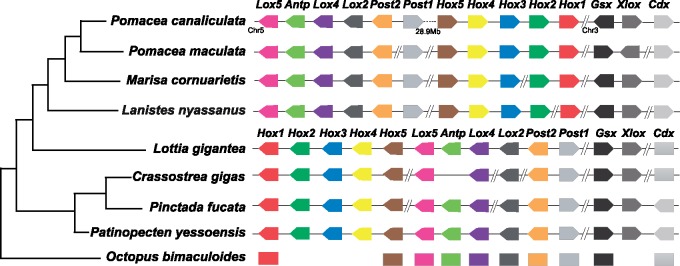
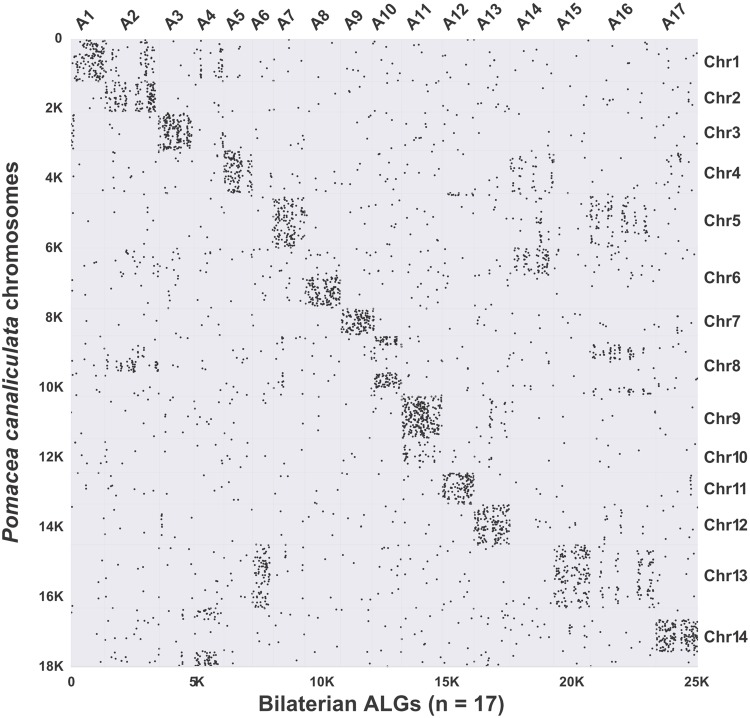
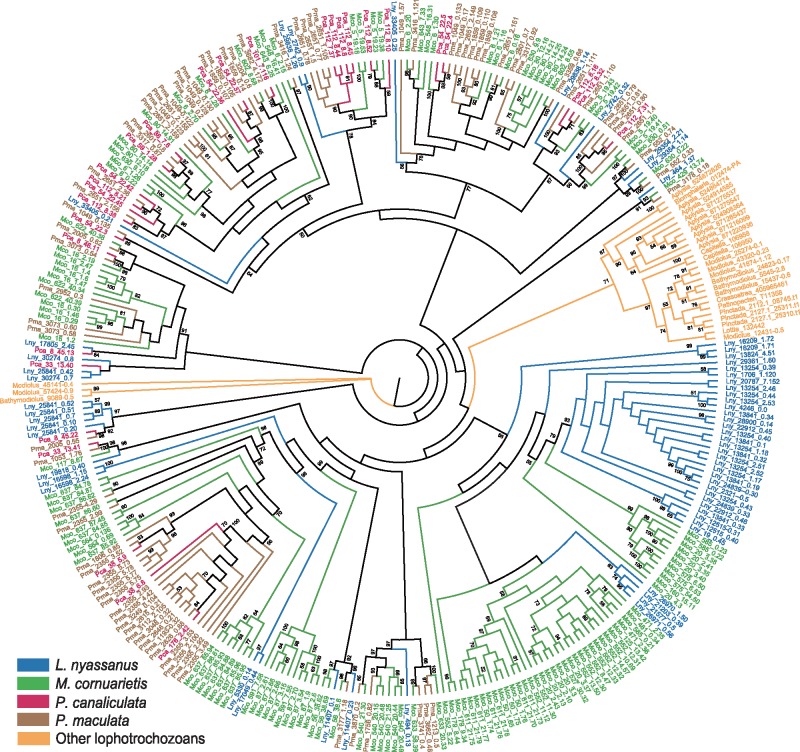
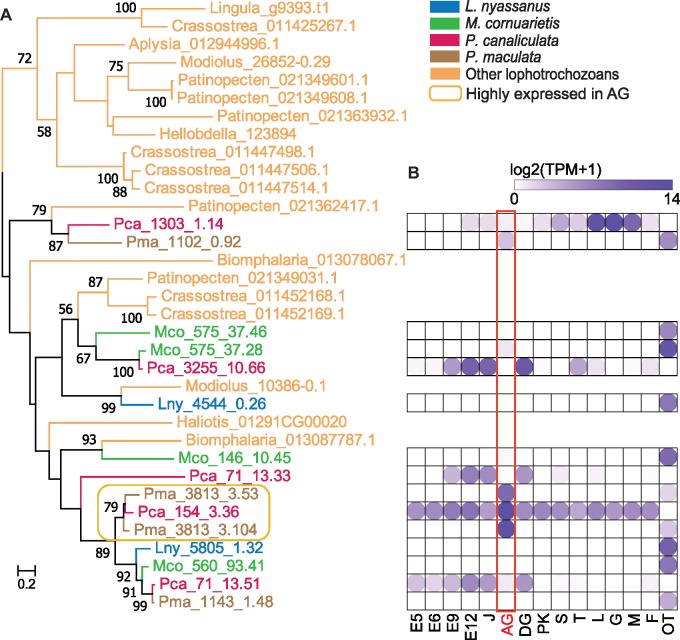
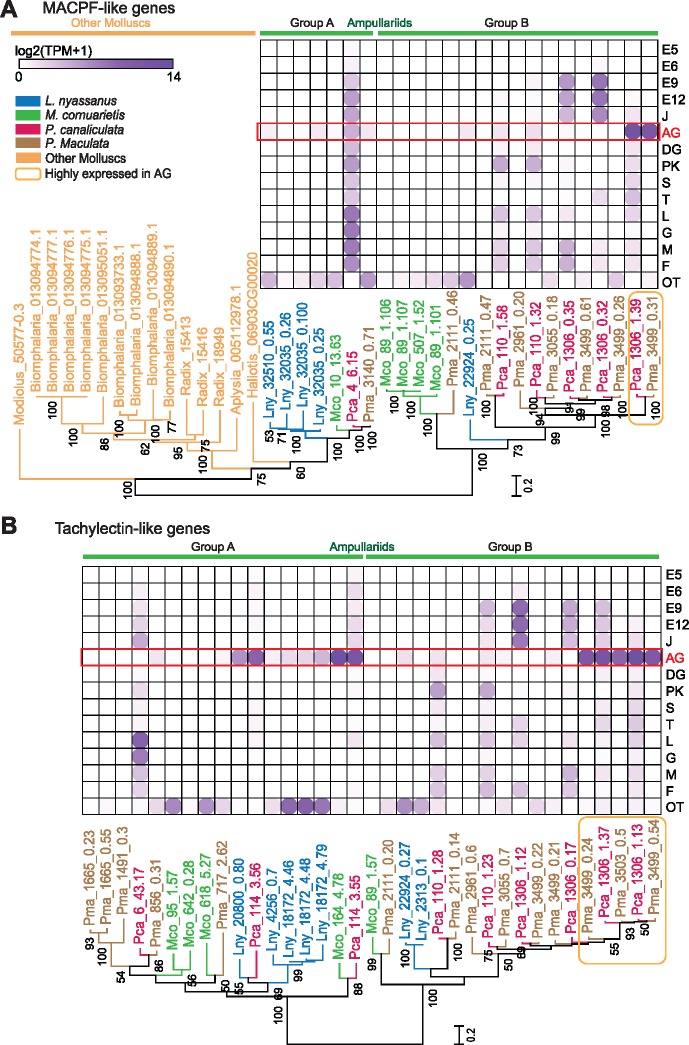
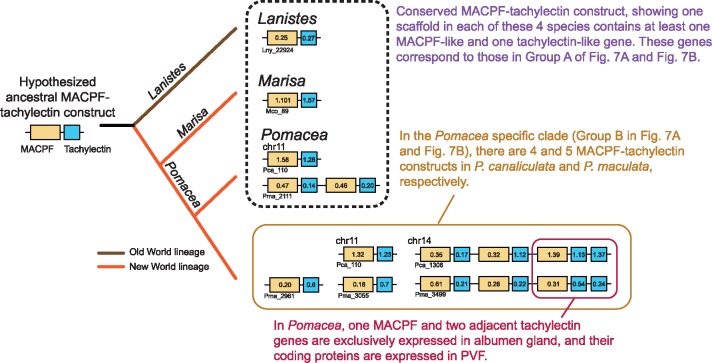
The family Ampullariidae includes both aquatic and amphibious apple snails. They are an emerging model for evolutionary studies due to the high diversity, ancient history, and wide geographical distribution. Insight into drivers of ampullariid evolution is hampered, however, by the lack of genomic resources. Here, we report the genomes of four ampullariids spanning the Old World (Lanistes nyassanus) and New World (Pomacea canaliculata, P. maculata, and Marisa cornuarietis) clades. The ampullariid genomes have conserved ancient bilaterial karyotype features and a novel Hox gene cluster rearrangement, making them valuable in comparative genomic studies. They have expanded gene families related to environmental sensing and cellulose digestion, which may have facilitated some ampullarids to become notorious invasive pests. In the amphibious Pomacea, novel acquisition of an egg neurotoxin and a protein for making the calcareous eggshell may have been key adaptations enabling their transition from underwater to terrestrial egg deposition.
Keywords: Hox genes; gastropod; gene duplication; genomics; interchromosome rearrangement; mollusc.
© The Author(s) 2019. Published by Oxford University Press on behalf of the Society for Molecular Biology and Evolution.
Figures
References
-
- Cadierno MP, Dreon MS, Heras H.. 2017. Apple snail perivitellin precursor properties help explain predators’ feeding behavior. Physiol Biochem Zool. 90(4):461–470. - PubMed
-
- Cowie RH. 2002. Apple snails (Ampullariidae) as agricultural pests: their biology, impacts and management In: Barker GM, editor. Molluscs as crop pests. Wallingford (United Kingdom: ): CAB International; p. 145–192.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
