

Homozygous Mutations in CSF1R Cause a Pediatric-Onset Leukoencephalopathy and Can Result in Congenital Absence of Microglia
- PMID: 30982608
- PMCID: PMC6506793
- DOI: 10.1016/j.ajhg.2019.03.010
Homozygous Mutations in CSF1R Cause a Pediatric-Onset Leukoencephalopathy and Can Result in Congenital Absence of Microglia
Abstract
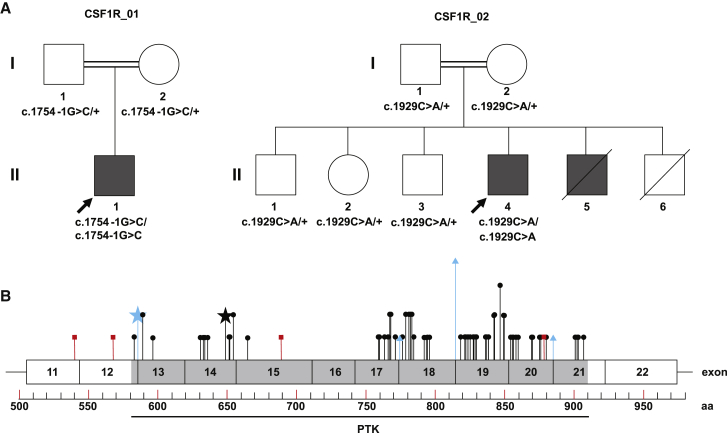
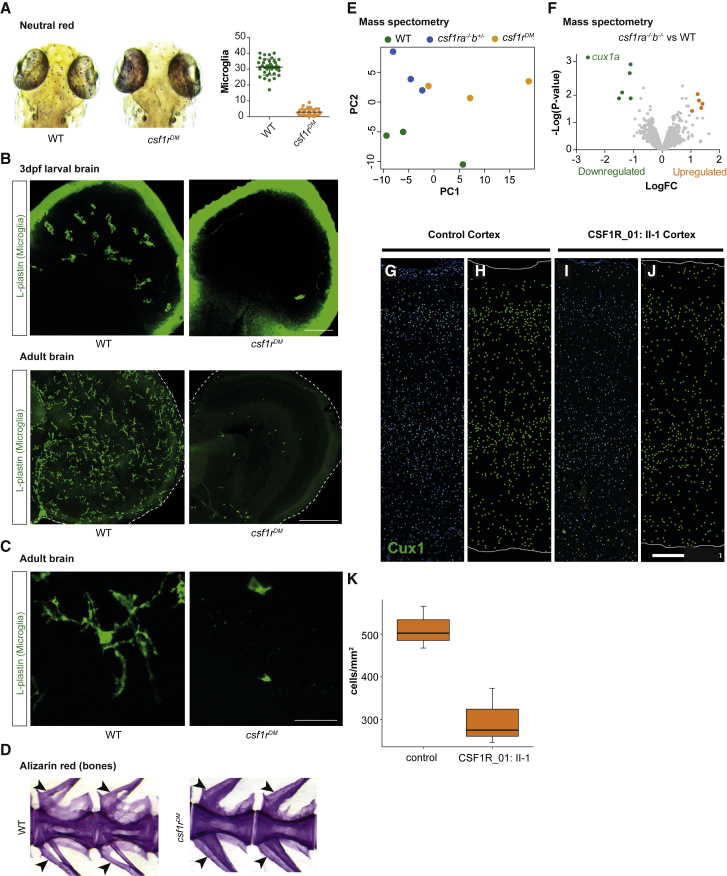
Microglia are CNS-resident macrophages that scavenge debris and regulate immune responses. Proliferation and development of macrophages, including microglia, requires Colony Stimulating Factor 1 Receptor (CSF1R), a gene previously associated with a dominant adult-onset neurological condition (adult-onset leukoencephalopathy with axonal spheroids and pigmented glia). Here, we report two unrelated individuals with homozygous CSF1R mutations whose presentation was distinct from ALSP. Post-mortem examination of an individual with a homozygous splice mutation (c.1754-1G>C) demonstrated several structural brain anomalies, including agenesis of corpus callosum. Immunostaining demonstrated almost complete absence of microglia within this brain, suggesting that it developed in the absence of microglia. The second individual had a homozygous missense mutation (c.1929C>A [p.His643Gln]) and presented with developmental delay and epilepsy in childhood. We analyzed a zebrafish model (csf1rDM) lacking Csf1r function and found that their brains also lacked microglia and had reduced levels of CUX1, a neuronal transcription factor. CUX1+ neurons were also reduced in sections of homozygous CSF1R mutant human brain, identifying an evolutionarily conserved role for CSF1R signaling in production or maintenance of CUX1+ neurons. Since a large fraction of CUX1+ neurons project callosal axons, we speculate that microglia deficiency may contribute to agenesis of the corpus callosum via reduction in CUX1+ neurons. Our results suggest that CSF1R is required for human brain development and establish the csf1rDM fish as a model for microgliopathies. In addition, our results exemplify an under-recognized form of phenotypic expansion, in which genes associated with well-recognized, dominant conditions produce different phenotypes when biallelically mutated.
Keywords: CSF1R; CUX1; agenesis corpus callosum; axonal spheroids; leukoencephalopathy; microglia; neuropathology; osteopetrosis; recessive; zebrafish.
Copyright © 2019 American Society of Human Genetics. Published by Elsevier Inc. All rights reserved.
Figures


References
-
- Paolicelli R.C., Bolasco G., Pagani F., Maggi L., Scianni M., Panzanelli P., Giustetto M., Ferreira T.A., Guiducci E., Dumas L. Synaptic pruning by microglia is necessary for normal brain development. Science. 2011;333:1456–1458. - PubMed
-
- Squarzoni P., Oller G., Hoeffel G., Pont-Lezica L., Rostaing P., Low D., Bessis A., Ginhoux F., Garel S. Microglia modulate wiring of the embryonic forebrain. Cell Rep. 2014;8:1271–1279. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
