

Attenuation of Novelty-Induced Hyperactivity of Gria1-/- Mice by Cannabidiol and Hippocampal Inhibitory Chemogenetics
- PMID: 30984001
- PMCID: PMC6449460
- DOI: 10.3389/fphar.2019.00309
Attenuation of Novelty-Induced Hyperactivity of Gria1-/- Mice by Cannabidiol and Hippocampal Inhibitory Chemogenetics
Abstract
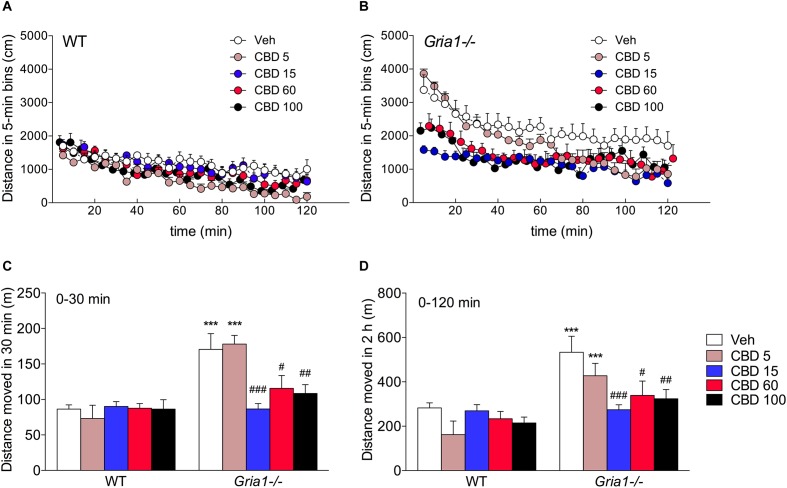
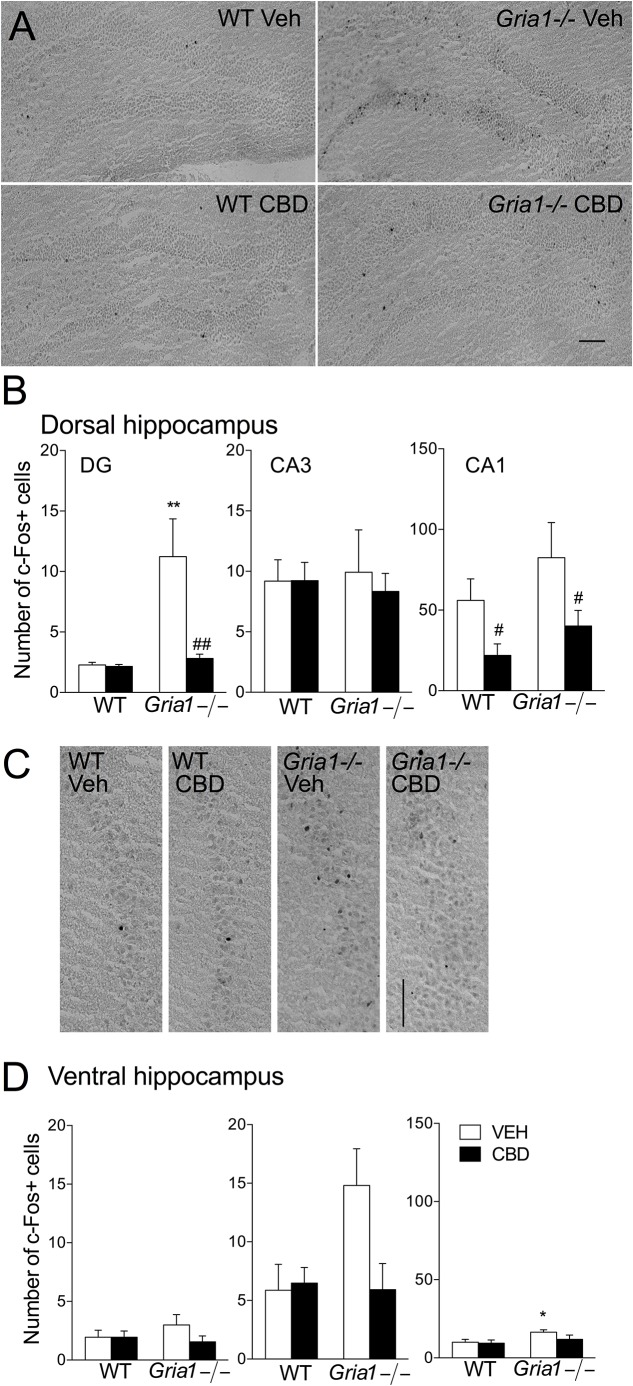
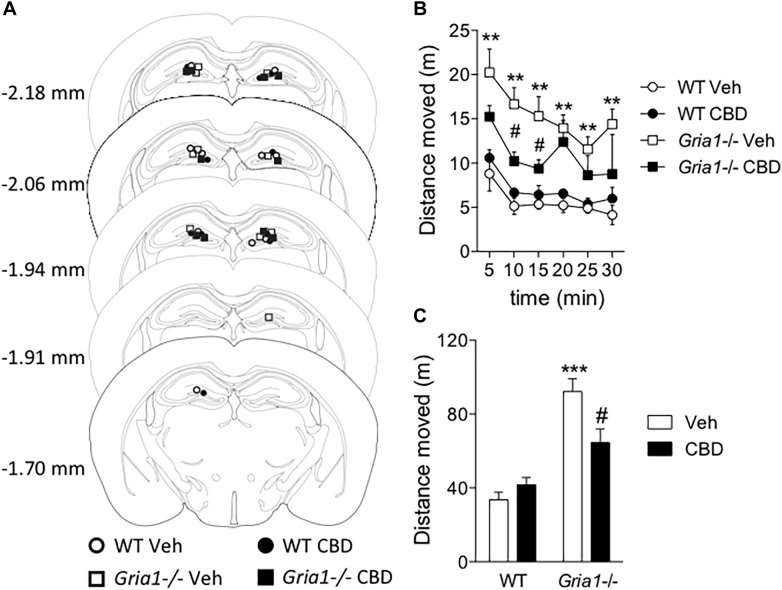
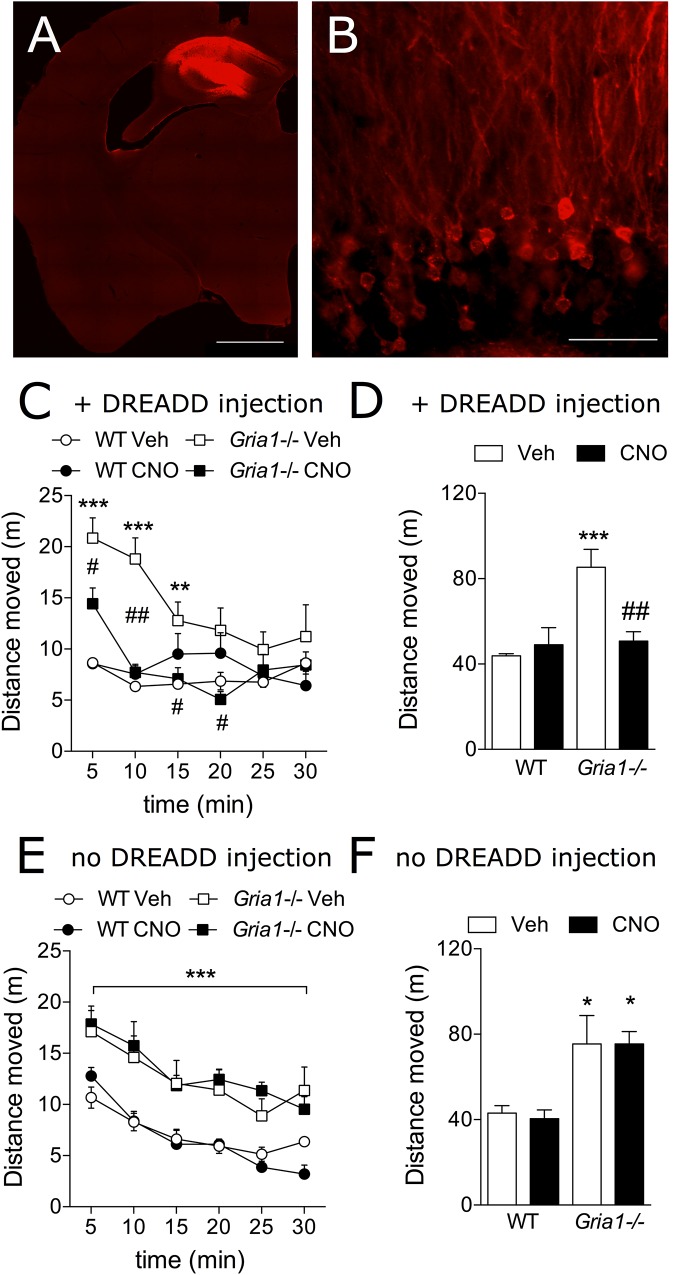
Gene-targeted mice with deficient AMPA receptor GluA1 subunits (Gria1-/- mice) show robust hyperlocomotion in a novel environment, suggesting them to constitute a model for hyperactivity disorders such as mania, schizophrenia and attention deficit hyperactivity disorder. This behavioral alteration has been associated with increased neuronal activation in the hippocampus, and it can be attenuated by chronic treatment with antimanic drugs, such as lithium, valproic acid, and lamotrigine. Now we found that systemic cannabidiol strongly blunted the hyperactivity and the hippocampal c-Fos expression of the Gria1-/- mice, while not affecting the wild-type littermate controls. Acute bilateral intra-dorsal hippocampal infusion of cannabidiol partially blocked the hyperactivity of the Gria1-/- mice, but had no effect on wild-types. The activation of the inhibitory DREADD receptor hM4Gi in the dorsal hippocampus by clozapine-N-oxide robustly inhibited the hyperactivity of the Gria1-/- mice, but had no effect on the locomotion of wild-type mice. Our results show that enhanced neuronal excitability in the hippocampus is associated with pronounced novelty-induced hyperactivity of GluA1 subunit-deficient mice. When this enhanced response of hippocampal neurons to novel stimuli is specifically reduced in the hippocampus by pharmacological treatment or by chemogenetic inhibition, Gria1-/- mice recover from behavioral hyperactivity, suggesting a hippocampal dysfunction in hyperactive behaviors that can be treated with cannabidiol.
Keywords: AMPA receptors; DREADD; c-Fos; cannabidiol; hM4Gi; hippocampus; hyperactivity; novelty.
Figures
References
-
- Barkus C., Feyder M., Graybeal C., Wright T., Wiedholz L., Izquierdo A., et al. (2011). Do GluA1 knockout mice exhibit behavioral abnormalities relevant to the negative or cognitive symptoms of schizophrenia and schizoaffective disorder? Neuropharmacology 62 1263–1272. 10.1016/j.neuropharm.2011.06.005 - DOI - PMC - PubMed
-
- Bianchi L., Ballini C., Colivicchi M. A., Della Corte L., Giovannini M. G., Pepeu G. (2003). Investigation on acetylcholine, aspartate, glutamate and GABA extracellular levels from ventral hippocampus during repeated exploratory activity in the rat. Neurochem. Res. 28 565–573. 10.1023/A:1022881625378 - DOI - PubMed
LinkOut - more resources
Full Text Sources
Molecular Biology Databases