

Milk Oligosaccharides From Different Cattle Breeds Influence Growth-Related Characteristics of Intestinal Cells
- PMID: 30984764
- PMCID: PMC6447651
- DOI: 10.3389/fnut.2019.00031
Milk Oligosaccharides From Different Cattle Breeds Influence Growth-Related Characteristics of Intestinal Cells
Abstract
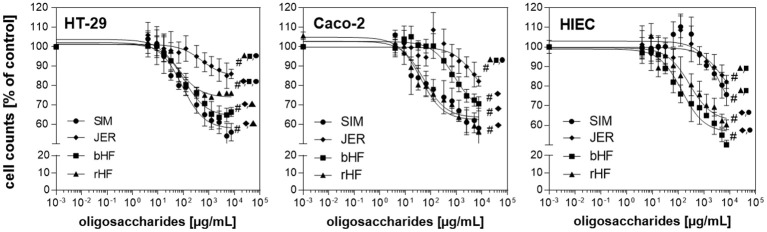
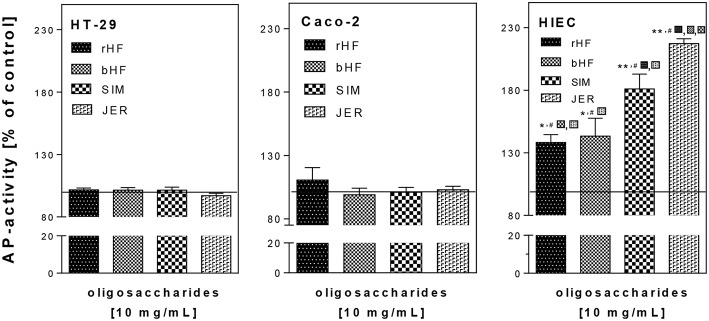
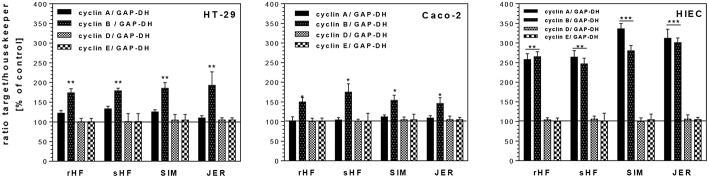
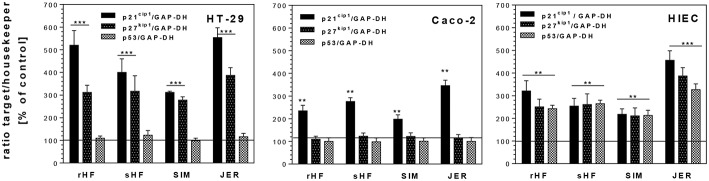
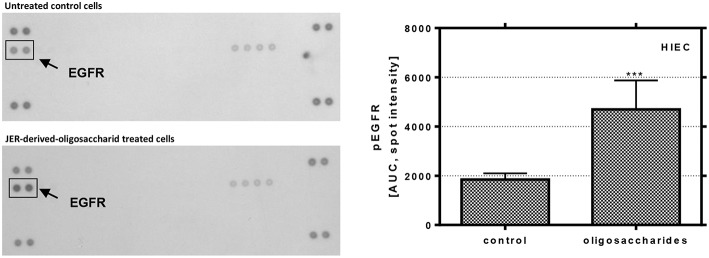
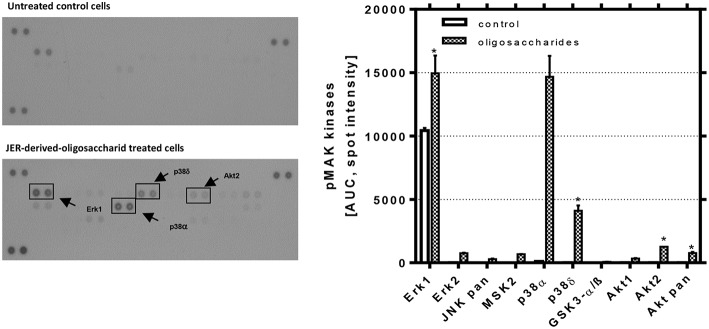
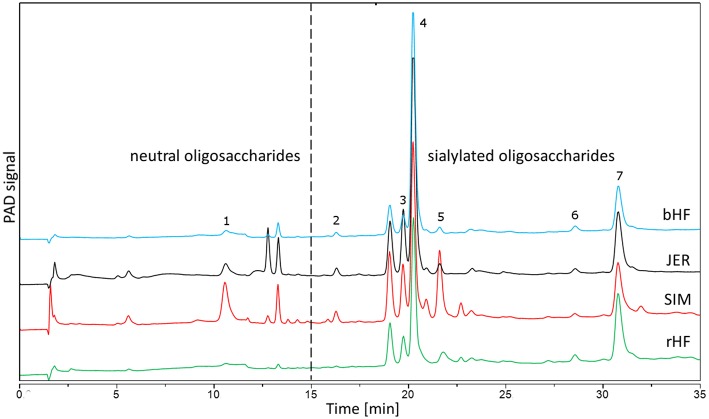
Oligosaccharides are present in human milk (HMO) in large amounts and in a high variety: Among other functions they are considered to influence the gut microbiota and gut maturation in infants. Due to the large volume of milk available bovine milk oligosaccharides (BMO) may be an alternative source of functional ingredients to potentially mimic HMO functions. Thus, we investigated direct effects of bovine milk oligosaccharides (BMO) from different cattle breeds on proliferation, differentiation and apoptosis in transformed (HT-29 and Caco-2) and non-transformed human intestinal cells (HIE cells). We observed a profound growth-inhibition effect induced by all BMO isolates in HT-29, Caco-2, and HIE cells in a dose-dependent manner. The effects varied not only between cell lines, i.e., HT-29 and Caco-2 cells were more sensitive than HIE cells, but also between the cattle breeds. Regarding the induction of differentiation, BMO induced differentiation only in HIE cells without affecting apoptosis. Cell cycle analysis via flow cytometry showed that growth inhibition was associated with a G2/M arrest in all cell lines. Expression levels detected by quantitative real-time RT-PCR revealed that this G2/M arrest was associated with changes in mRNA expression levels of cyclin A and B. Cyclin-dependent kinase inhibitors p21 cip1 and p27 kip1 and the tumor suppressor p53 were only enhanced in HIE cells necessary for arresting cells in the G2/M phase and induction of differentiation. In HT-29 and Caco-2 cells, a loss of p53 expression failed to induce G2/M associated induction of differentiation. The HIE cell specific differentiation induced by BMO was a result of influencing the phosphorylation states of EGFR (epidermal growth factor receptor) and MAP kinase, i.e., ERK1/2 (extracellular signal-regulated kinase 1/2), p38-α, and Akt2 phosphorylation. These results suggest that BMO inhibited intestinal cell proliferation and altered cell cycle dynamics by affecting corresponding regulator genes and mitogen-activated protein kinase signaling. As the development and maturation of digestive and absorptive processes depends on gut differentiation processes, our in vitro experiments show that breed-specific BMO are natural substances influencing various parameter which may be important in vivo in gastrointestinal development. This, however, needs to be proven in future studies.
Keywords: BMO; EGFR-ERK signaling; bovine milk oligosaccharides; cell cycle dynamics; differentiation; interbreed variation.
Figures
References
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous