

Birth and Resuscitation of (p)ppGpp Induced Antibiotic Tolerant Persister Cells
- PMID: 30988388
- PMCID: PMC6465370
- DOI: 10.1038/s41598-019-42403-7
Birth and Resuscitation of (p)ppGpp Induced Antibiotic Tolerant Persister Cells
Abstract
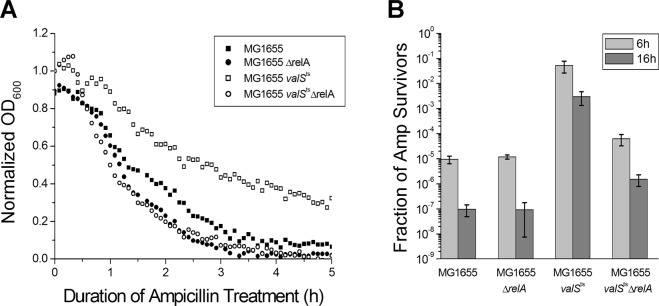
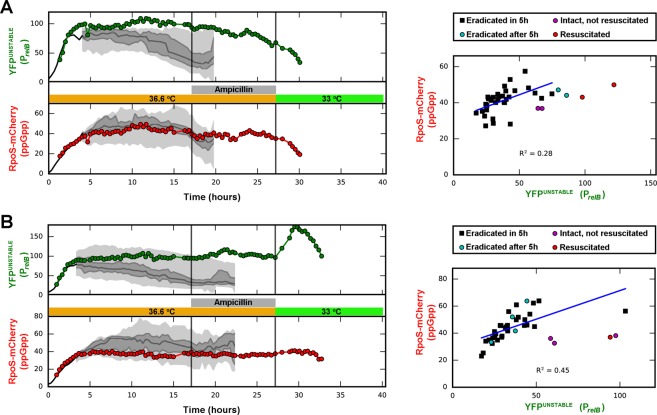
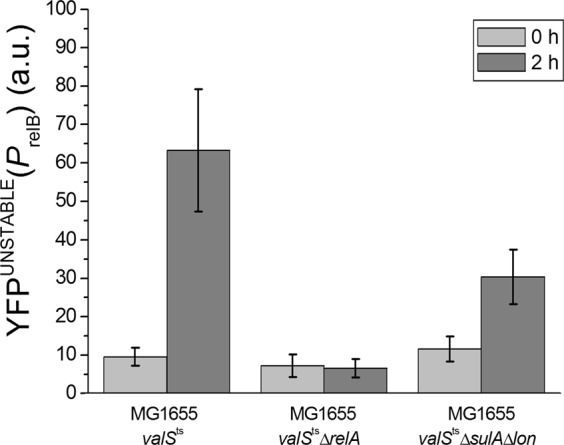
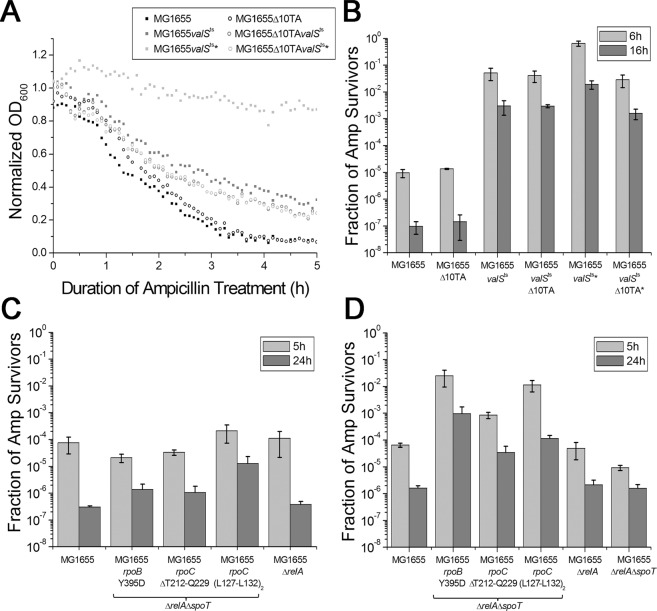
Transient antibiotic treatment typically eradicates most sensitive bacteria except a few survivors called persisters. The second messenger (p)ppGpp plays a key role in persister formation in Escherichia coli populations but the underlying mechanisms have remained elusive. In this study we induced (p)ppGpp synthesis by modulating tRNA charging and then directly observed the stochastic appearance, antibiotic tolerance, and resuscitation of persister cells using live microscopy. Different physiological parameters of persister cells as well as their regularly growing ancestors and sisters were continuously monitored using fluorescent reporters. Our results confirmed previous findings that high (p)ppGpp levels are critical for persister formation, but the phenomenon remained strikingly stochastic without any correlation between (p)ppGpp levels and antibiotic tolerance on the single-cell level. We could not confirm previous notions that persisters exhibit markedly low concentrations of intracellular ATP or were linked to post-transcriptional effects of (p)ppGpp through the activation of small genetic elements known as toxin-antitoxin (TA) modules. Instead, we suggest that persister cell formation under regular conditions is driven by the transcriptional response to increased (p)ppGpp levels.
Conflict of interest statement
The authors declare no competing interests.
Figures


References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
