

Impaired PSII proteostasis triggers a UPR-like response in the var2 mutant of Arabidopsis
- PMID: 30989223
- PMCID: PMC6598079
- DOI: 10.1093/jxb/erz151
Impaired PSII proteostasis triggers a UPR-like response in the var2 mutant of Arabidopsis
Abstract
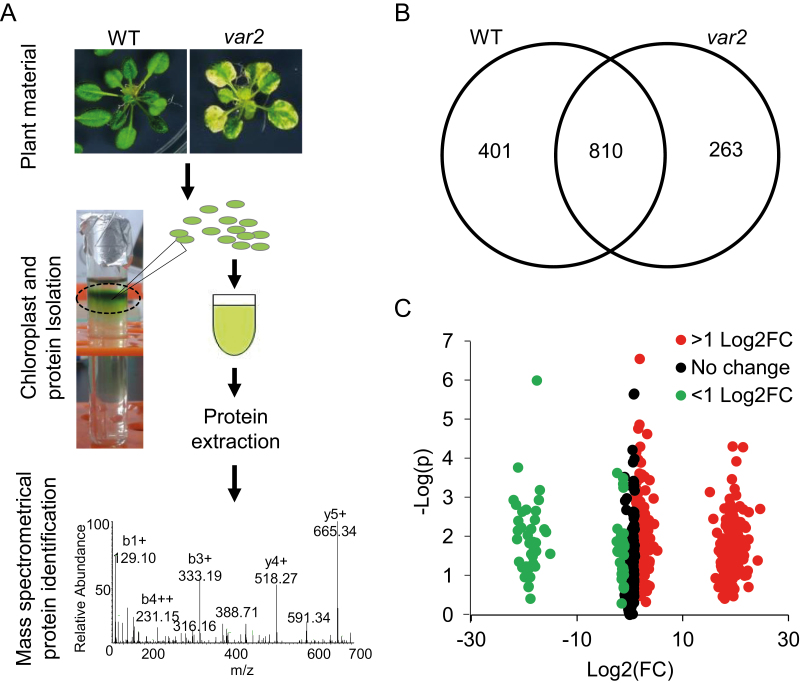
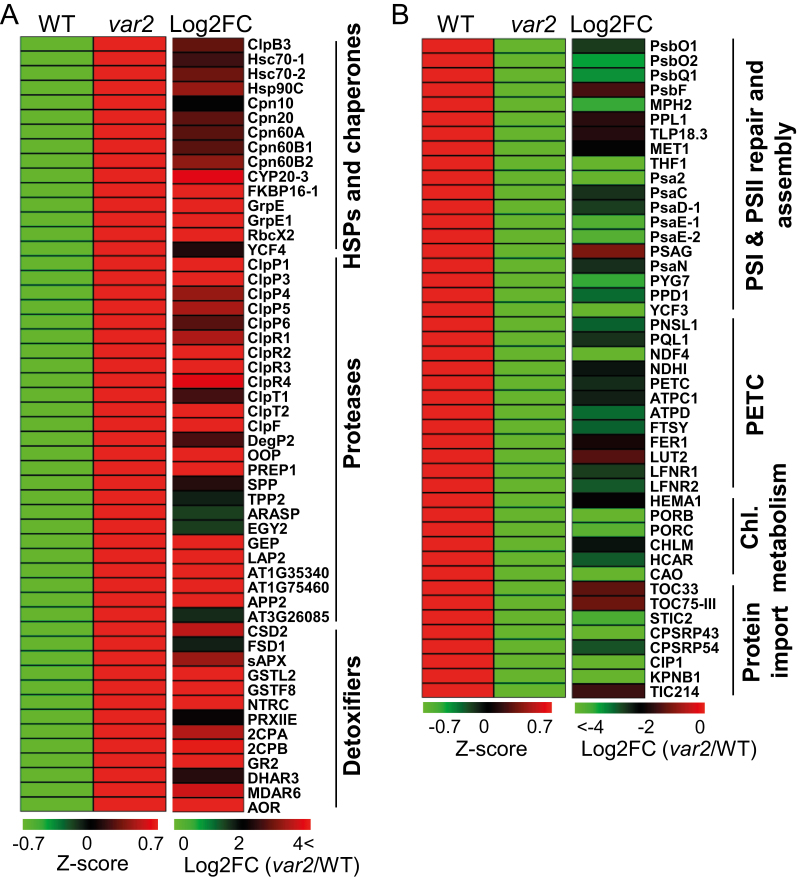
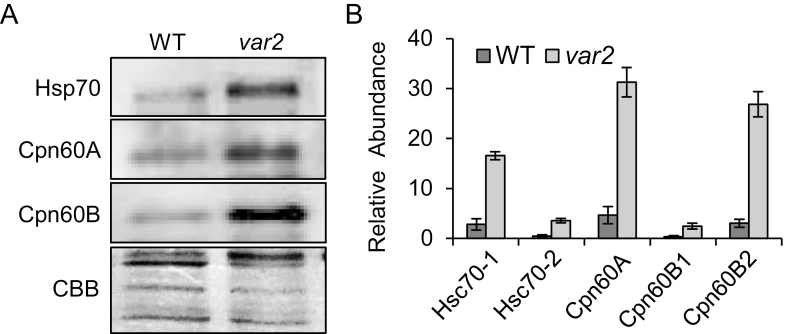
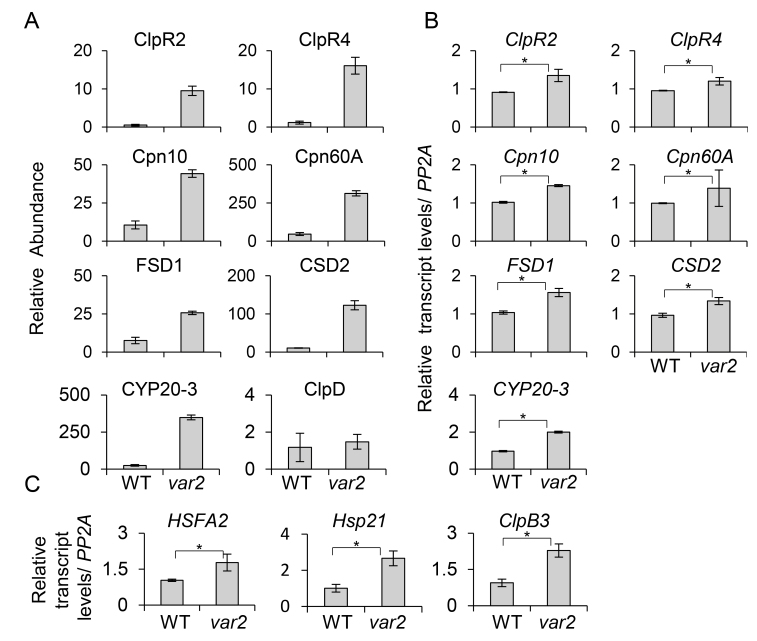
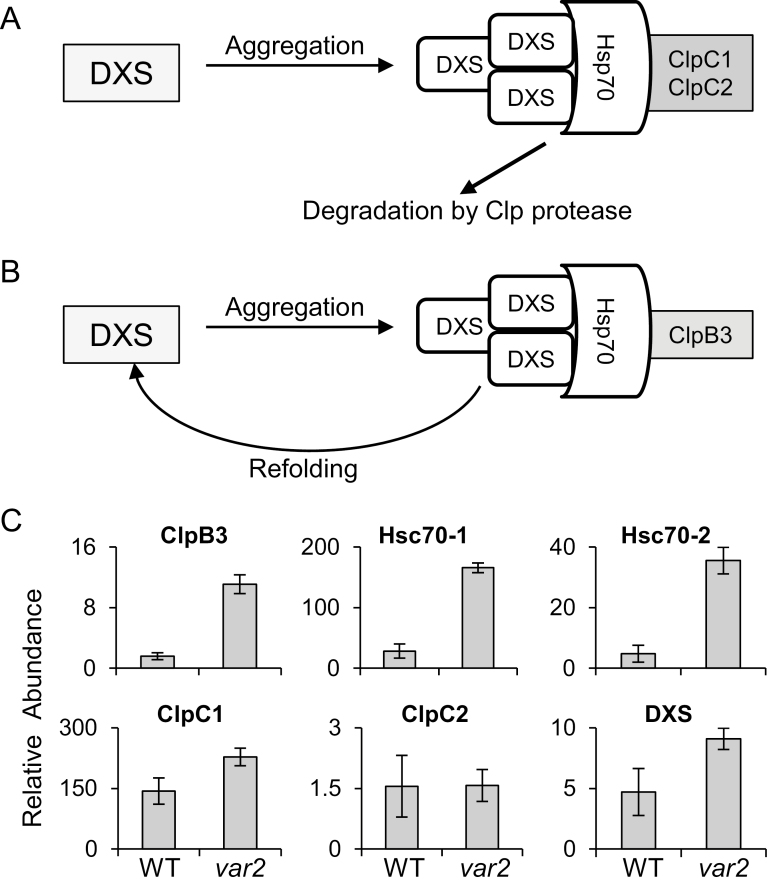
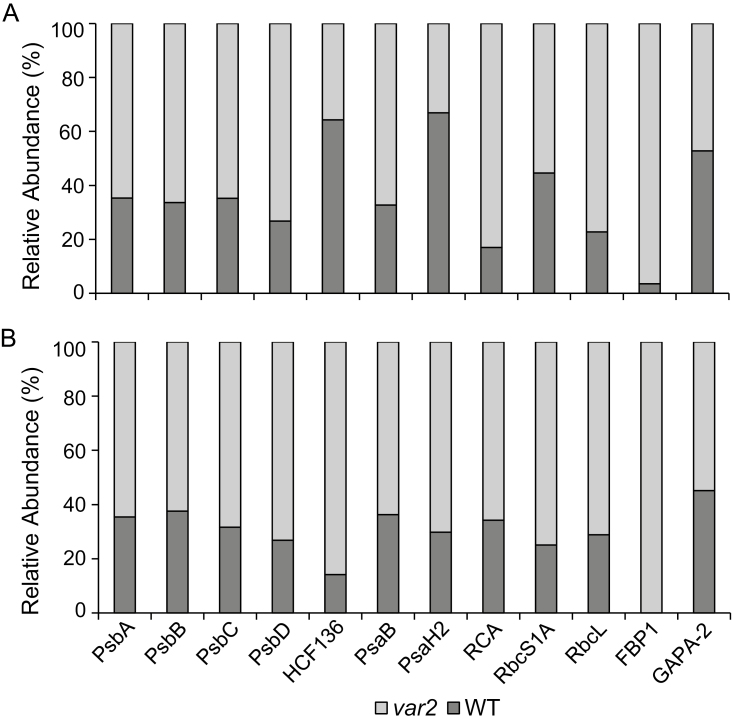
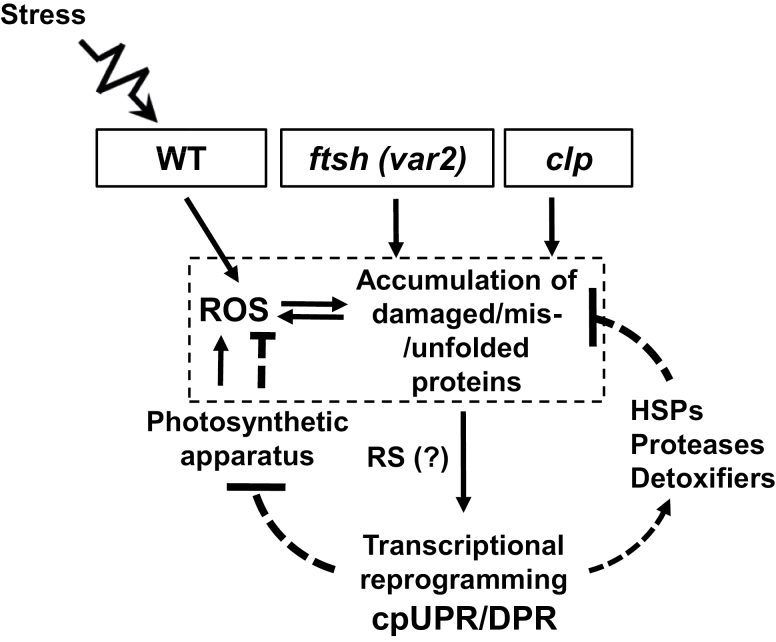
Cellular protein homeostasis (proteostasis) is maintained through the balance between de novo synthesis and proteolysis. The unfolded/misfolded protein response (UPR) that is triggered by stressed endoplasmic reticulum (ER) also plays an important role in proteostasis in both plants and animals. Although ER-triggered UPR has been extensively studied in plants, the molecular mechanisms underlying mitochondrial and chloroplastic UPRs are largely uncharacterized despite the fact that these organelles are sites of production of harmful reactive oxygen species (ROS), which damage proteins. In this study, we demonstrate that chloroplasts of the Arabidopsis yellow leaf variegation 2 (var2) mutant, which lacks the metalloprotease FtsH2, accumulate damaged chloroplast proteins and trigger a UPR-like response, namely the accumulation of a suite of chloroplast proteins involved in protein quality control (PQC). These PQC proteins include heat-shock proteins, chaperones, proteases, and ROS detoxifiers. Given that FtsH2 functions primarily in photosystem II proteostasis, the accumulation of PQC-related proteins may balance the FtsH2 deficiency. Moreover, the apparent up-regulation of the cognate transcripts indicates that the accumulation of PQC-related proteins in var2 is probably mediated by retrograde signaling, indicating the occurrence of a UPR-like response in var2.
Keywords: PSII repair; Photosystem II; damaged protein response; proteostasis; retrograde signaling; singlet oxygen; unfolded protein response.
© The Author(s) 2019. Published by Oxford University Press on behalf of the Society for Experimental Biology.
Figures
References
-
- Adam Z, Rudella A, van Wijk KJ. 2006. Recent advances in the study of Clp, FtsH and other proteases located in chloroplasts. Current Opinion in Plant Biology 9, 234–240. - PubMed
-
- Andersson B, Anderson JM. 1980. Lateral heterogeneity in the distribution of chlorophyll-protein complexes of the thylakoid membranes of spinach chloroplasts. Biochimica et Biophysica Acta 593, 427–440. - PubMed
-
- Apel K, Hirt H. 2004. Reactive oxygen species: metabolism, oxidative stress, and signal transduction. Annual Review of Plant Biology 55, 373–399. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
