

Experimental Evidence that Stochasticity Contributes to Bacterial Composition and Functioning in a Decomposer Community
- PMID: 30992354
- PMCID: PMC6469972
- DOI: 10.1128/mBio.00568-19
Experimental Evidence that Stochasticity Contributes to Bacterial Composition and Functioning in a Decomposer Community
Abstract
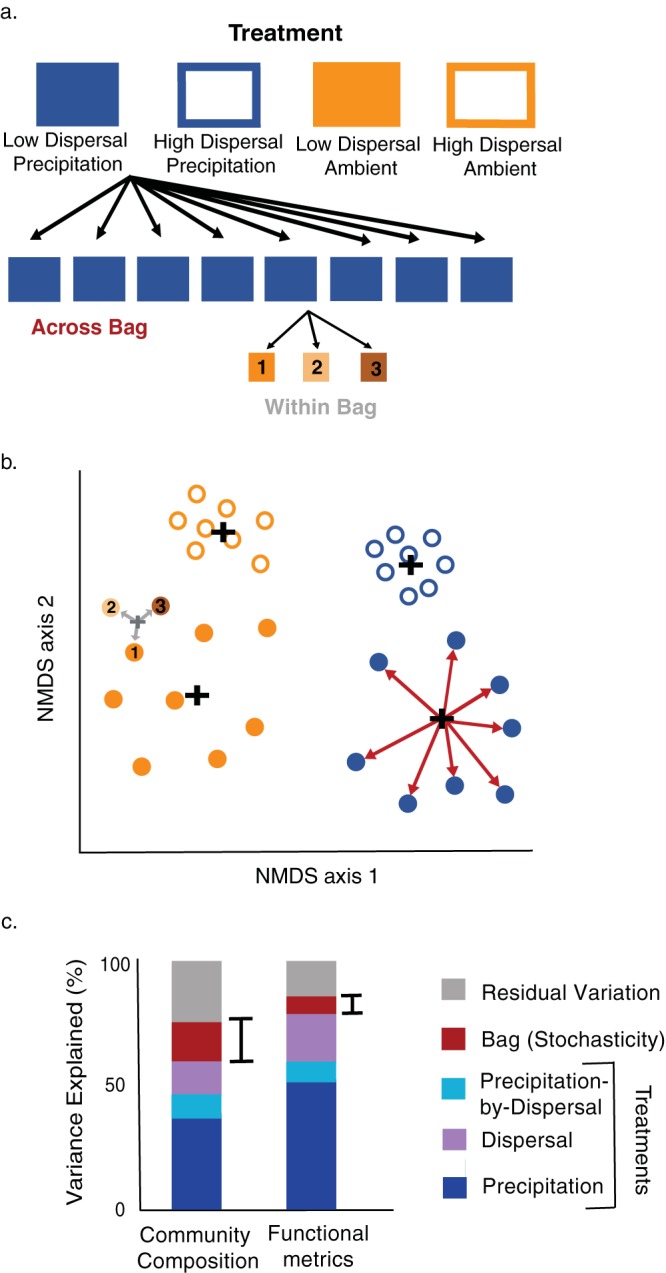
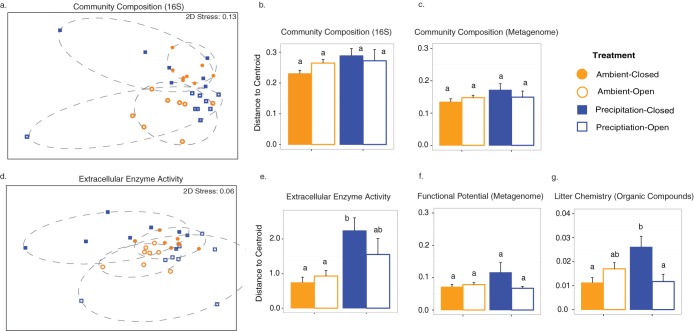
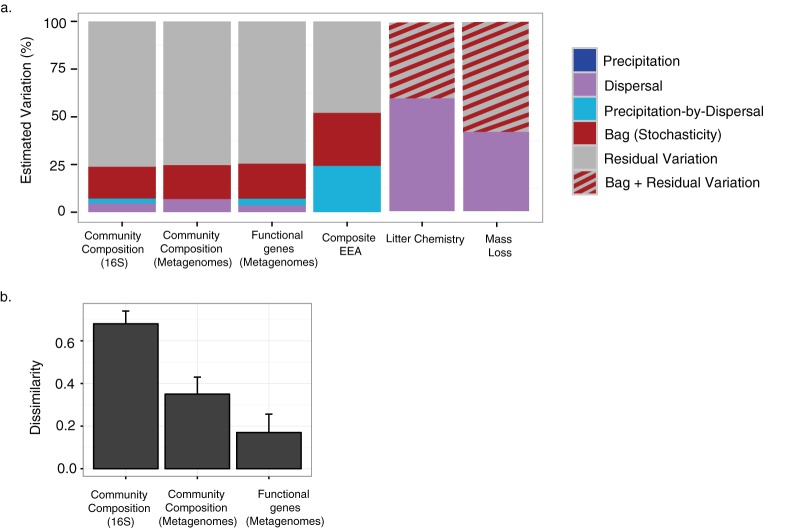
Stochasticity emerging from random differences in replication, death, mutation, and dispersal is thought to alter the composition of ecological communities. However, the importance of stochastic effects remains somewhat speculative because stochasticity is not directly measured but is instead inferred from unexplained variations in beta-diversity. Here, we performed a field experiment to more directly disentangle the role of stochastic processes, environmental selection, and dispersal in the composition and functioning of a natural bacterial decomposer community in the field. To increase our ability to detect these effects, we reduced initial biological and environmental heterogeneity using replicate nylon litterbags in the field. We then applied two treatments: ambient/added precipitation and bacterial and fungal dispersal using "open" litterbags (made from 18.0-μm-pore-size mesh) ("open bacterial dispersal") versus bacterial and fungal dispersal using "closed" litterbags (made from 22.0-μm-pore-size mesh) ("closed bacterial dispersal"). After 5 months, we assayed composition and functioning by the use of three subsamples from each litterbag to disentangle stochastic effects from residual variation. Our results indicate that stochasticity via ecological drift can contribute to beta-diversity in bacterial communities. However, residual variation, which had previously been included in stochasticity estimates, accounted for more than four times as much variability. At the same time, stochastic effects on beta-diversity were not attenuated at the functional level, as measured by genetic functional potential and extracellular enzyme activity. Finally, dispersal was found to interact with precipitation availability to influence the degree to which stochasticity contributed to functional variation. Together, our results demonstrate that the ability to quantify stochastic processes is key to understanding microbial diversity and its role in ecosystem functioning.IMPORTANCE Randomness can alter the diversity and composition of ecological communities. Such stochasticity may therefore obscure the relationship between the environment and community composition and hinder our ability to predict the relationship between biodiversity and ecosystem functioning. This study investigated the role of stochastic processes, environmental selection, and dispersal in microbial composition and its functioning on an intact field community. To do this, we used a controlled and replicated experiment that was similar to that used to study population genetics in the laboratory. Our study showed that, while the stochastic effects on taxonomic composition are smaller than expected, the degree to which stochasticity contributes to variability in ecosystem processes may be much higher than previously assumed.
Keywords: beta-diversity; dispersal; ecological drift; ecosystem functioning; stochastic.
Copyright © 2019 Albright et al.
Figures
References
-
- Hubbell SP. 2001. The unified neutral theory of biodiversity and biogeography. Princeton Univeristy Press, Princeton, NJ. - PubMed
