

Cardiomyocyte glucocorticoid and mineralocorticoid receptors directly and antagonistically regulate heart disease in mice
- PMID: 30992401
- PMCID: PMC7082727
- DOI: 10.1126/scisignal.aau9685
Cardiomyocyte glucocorticoid and mineralocorticoid receptors directly and antagonistically regulate heart disease in mice
Abstract
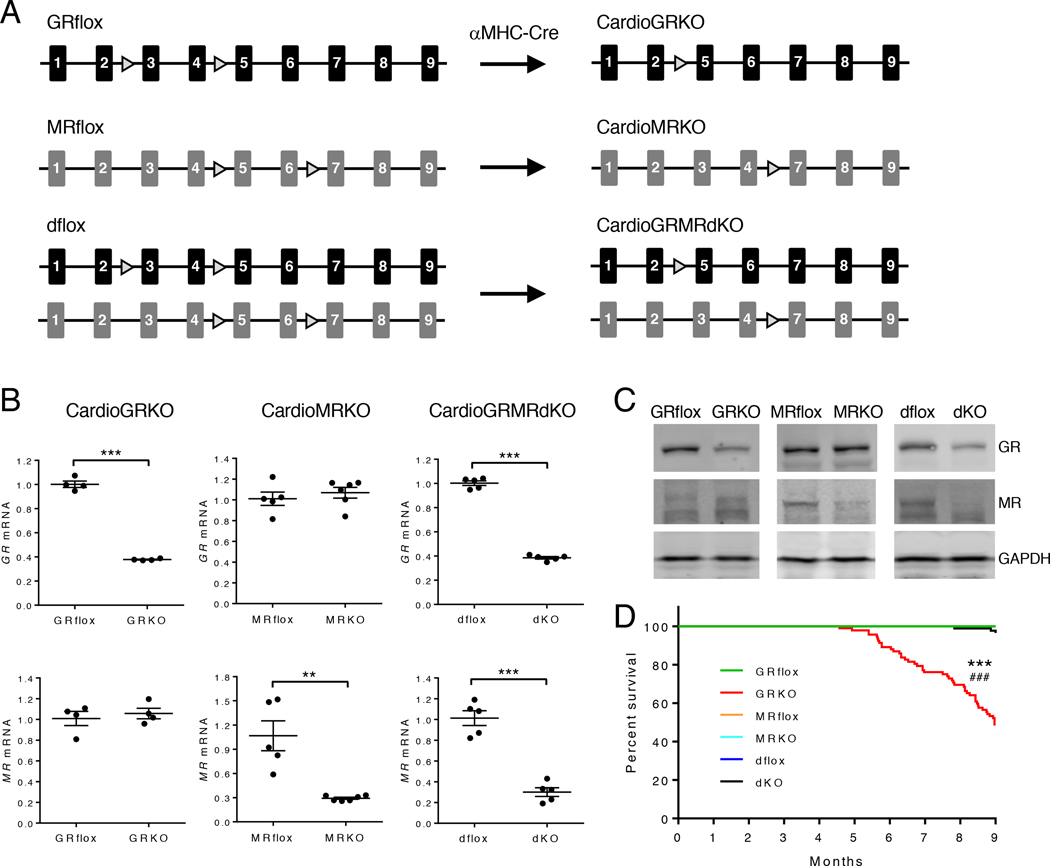
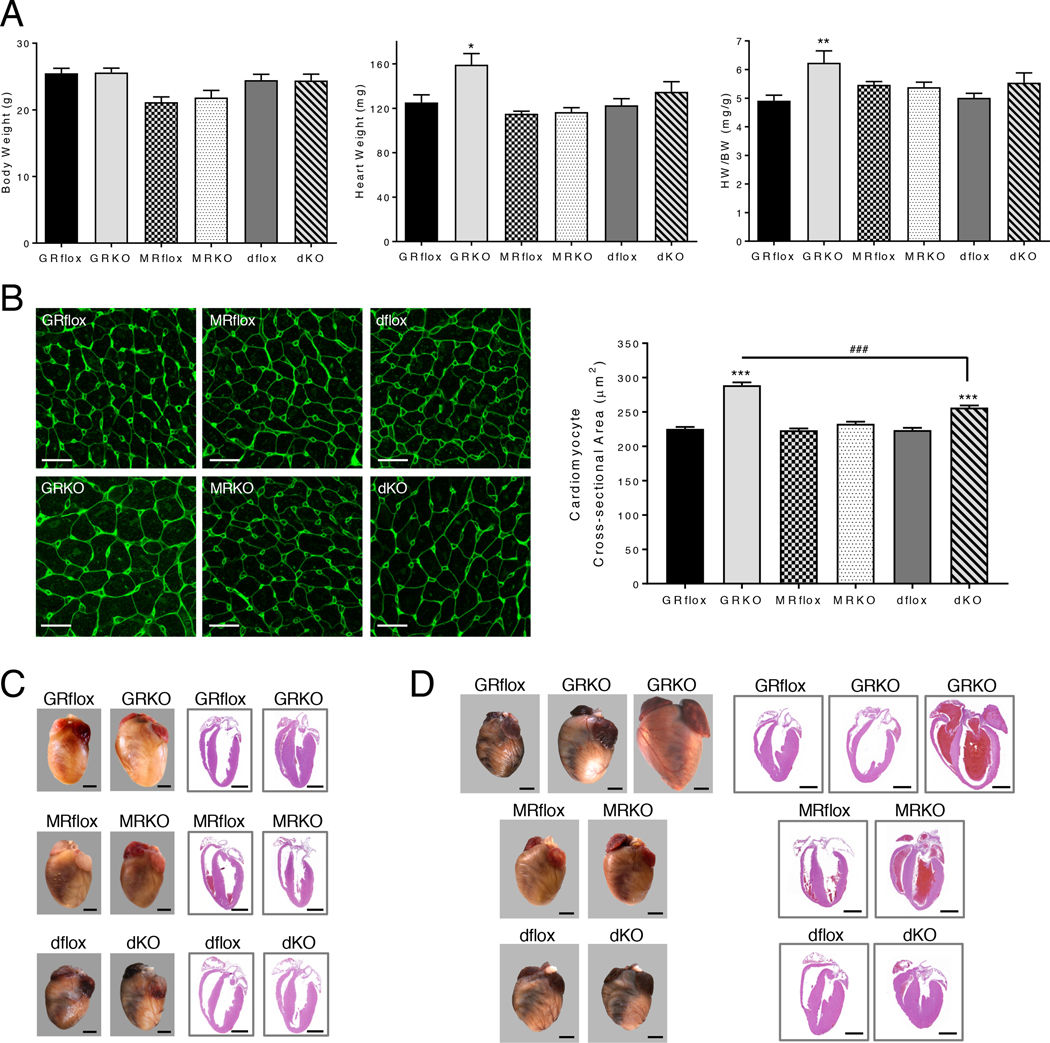
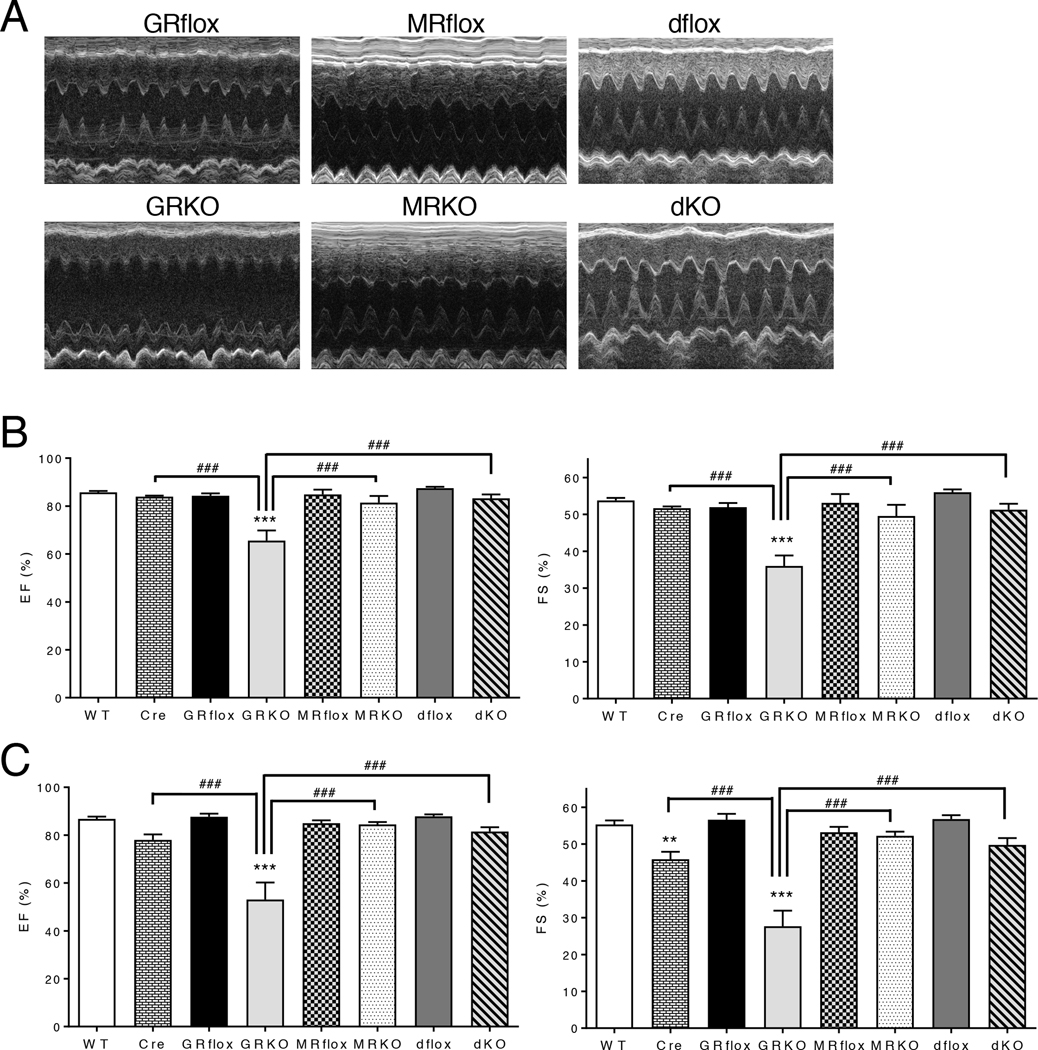
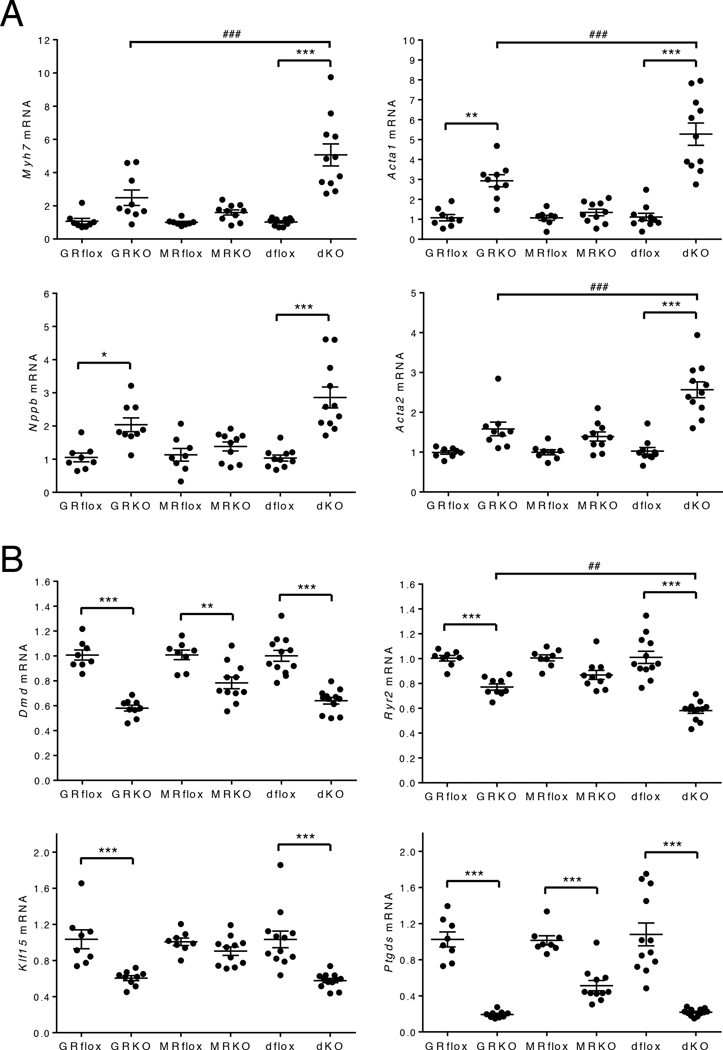
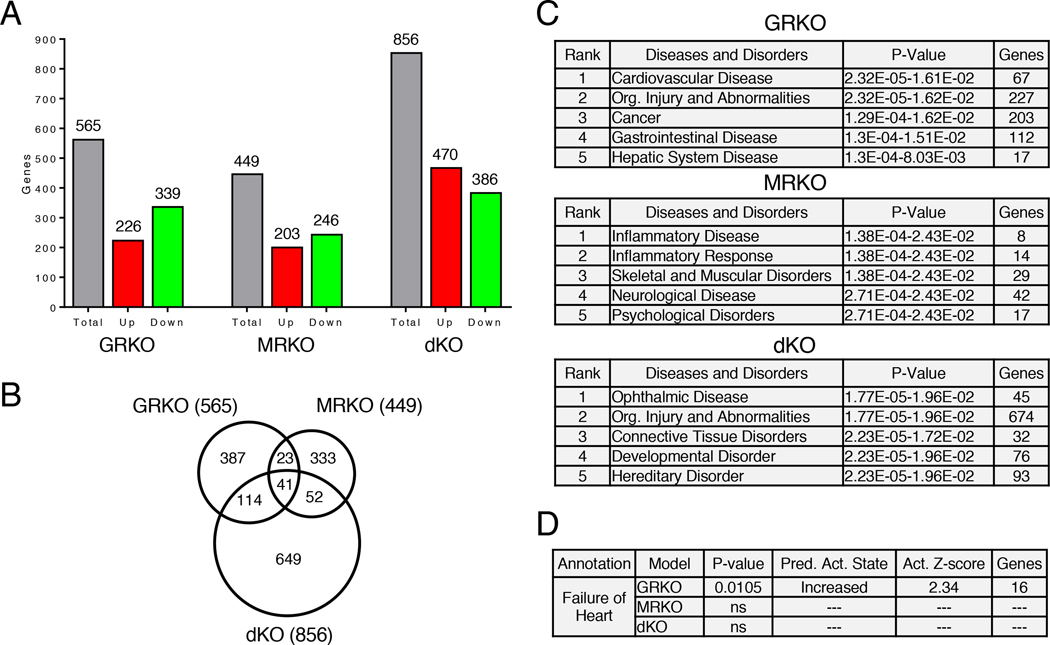
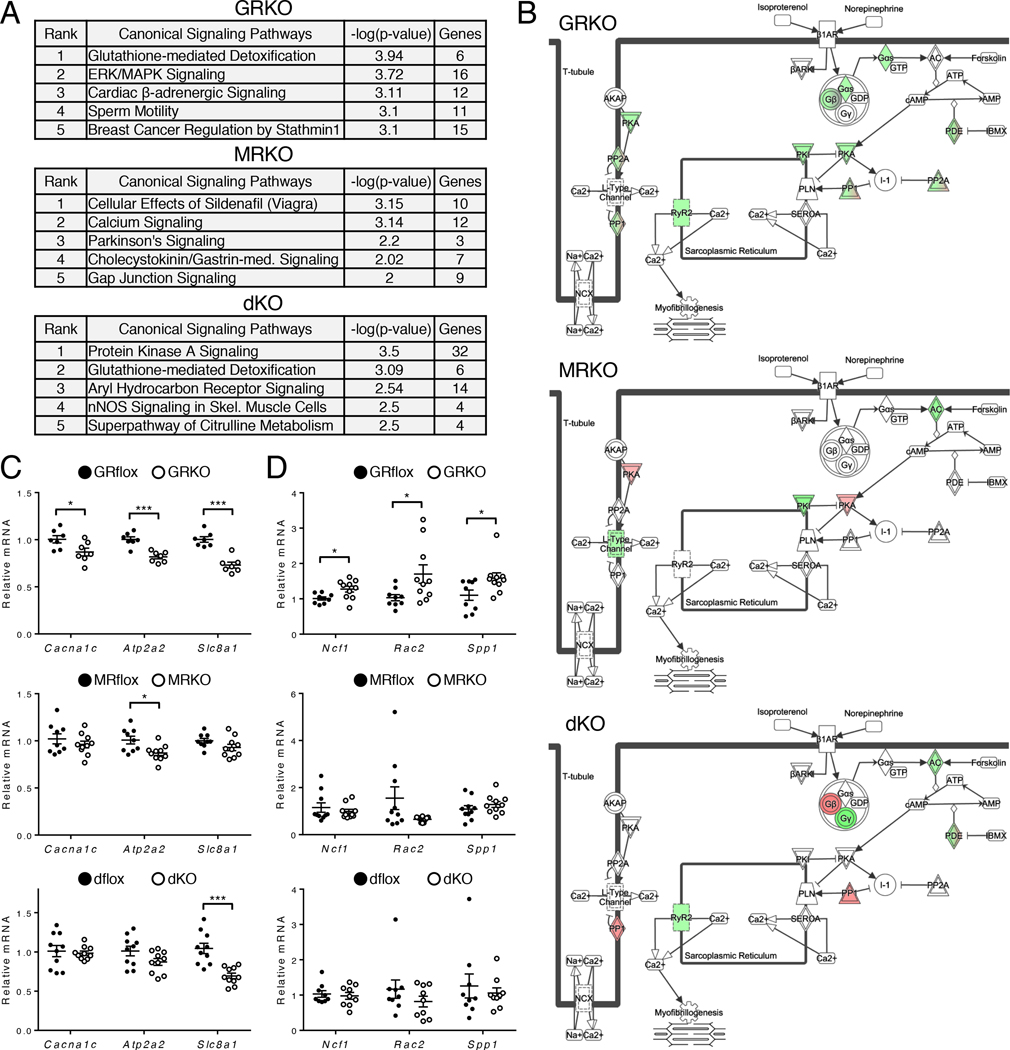
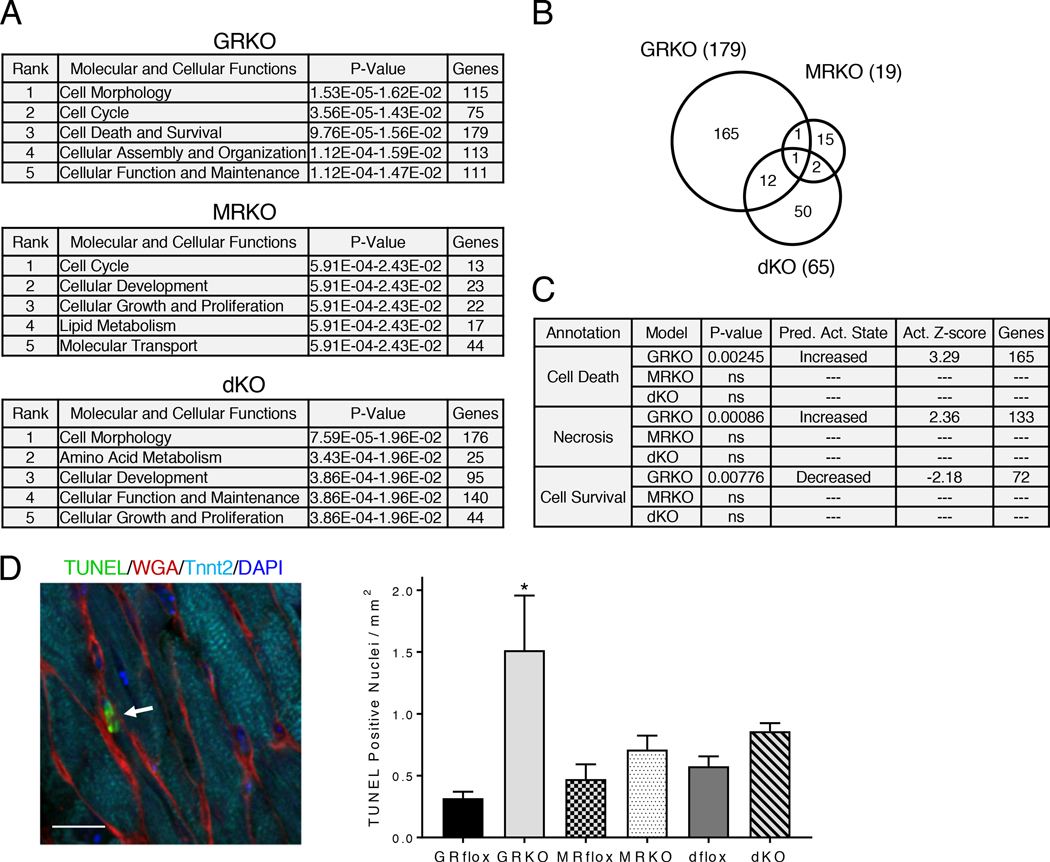
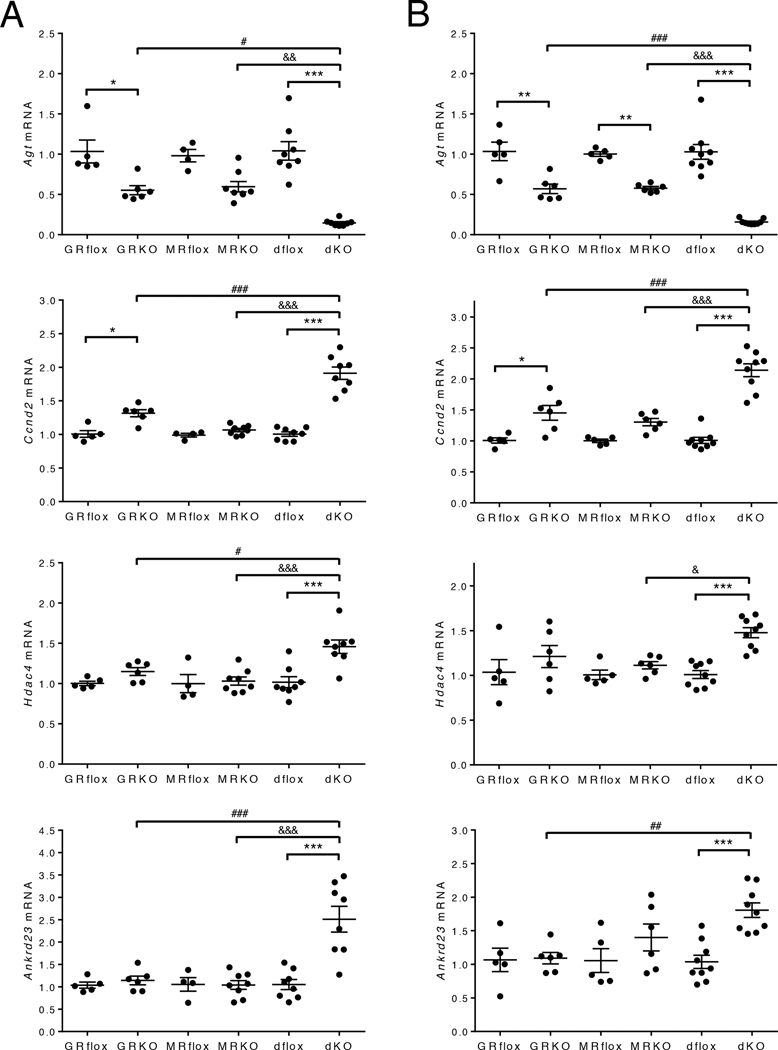
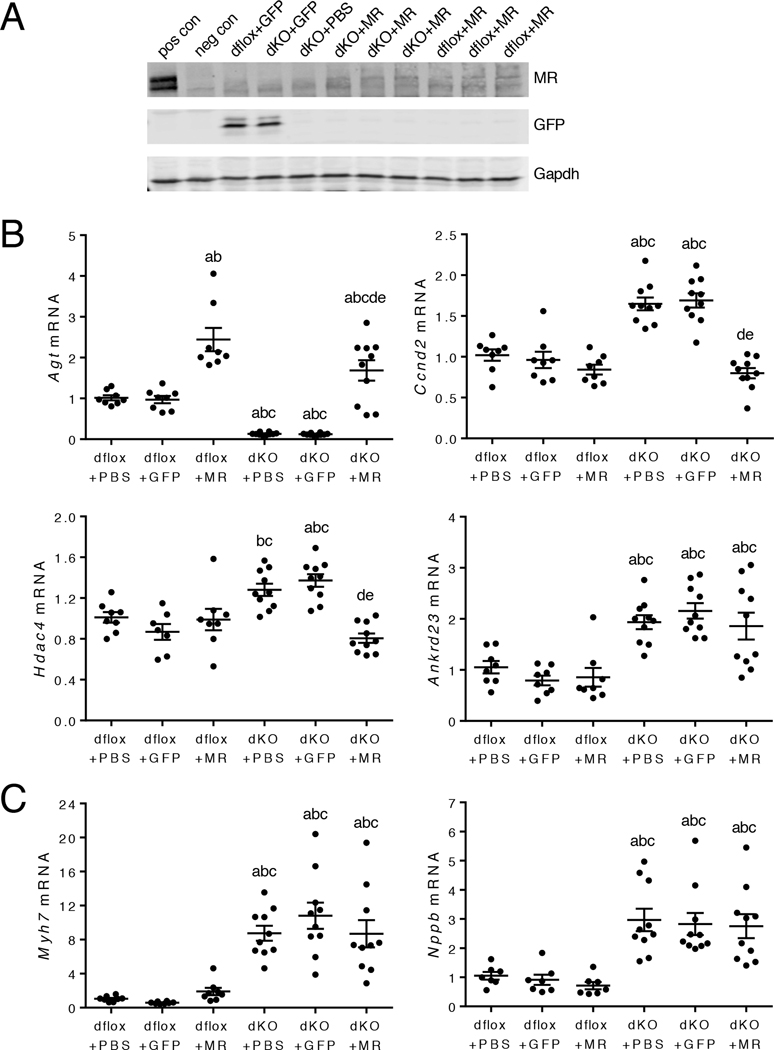
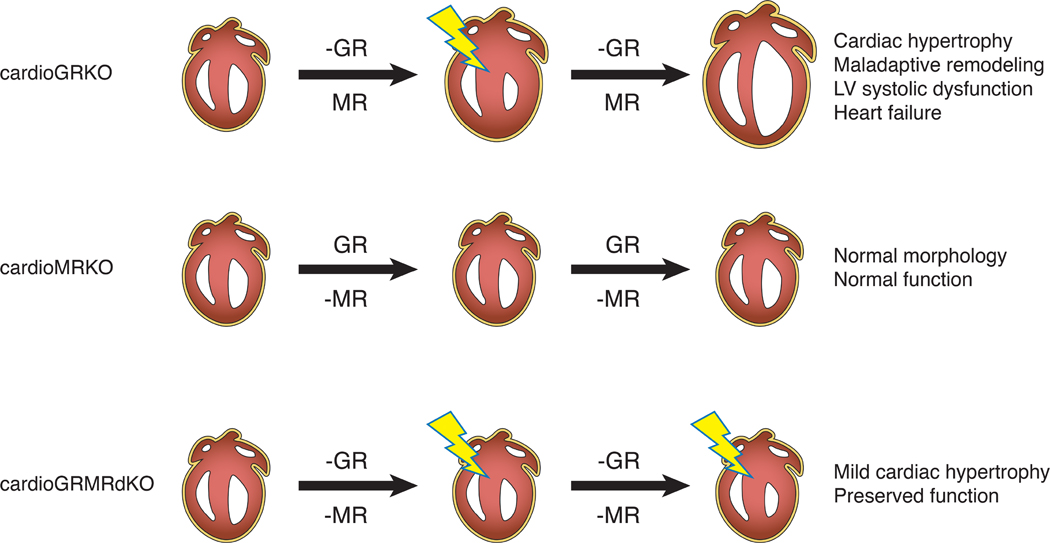
Stress is increasingly associated with heart dysfunction and is linked to higher mortality rates in patients with cardiometabolic disease. Glucocorticoids are primary stress hormones that regulate homeostasis through two nuclear receptors, the glucocorticoid receptor (GR) and mineralocorticoid receptor (MR), both of which are present in cardiomyocytes. To examine the specific and coordinated roles that these receptors play in mediating the direct effects of stress on the heart, we generated mice with cardiomyocyte-specific deletion of GR (cardioGRKO), MR (cardioMRKO), or both GR and MR (cardioGRMRdKO). The cardioGRKO mice spontaneously developed cardiac hypertrophy and left ventricular systolic dysfunction and died prematurely from heart failure. In contrast, the cardioMRKO mice exhibited normal heart morphology and function. Despite the presence of myocardial stress, the cardioGRMRdKO mice were resistant to the cardiac remodeling, left ventricular dysfunction, and early death observed in the cardioGRKO mice. Gene expression analysis revealed the loss of gene changes associated with impaired Ca2+ handling, increased oxidative stress, and enhanced cell death and the presence of gene changes that limited the hypertrophic response and promoted cardiomyocyte survival in the double knockout hearts. Reexpression of MR in cardioGRMRdKO hearts reversed many of the cardioprotective gene changes and resulted in cardiac failure. These findings reveal a critical role for balanced cardiomyocyte GR and MR stress signaling in cardiovascular health. Therapies that shift stress signaling in the heart to favor more GR and less MR activity may provide an improved approach for treating heart disease.
Copyright © 2019 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Figures
Comment in
-
Balancing stress signalling in the heart.Nat Rev Cardiol. 2019 Jul;16(7):384-385. doi: 10.1038/s41569-019-0207-0. Nat Rev Cardiol. 2019. PMID: 31068688 No abstract available.
References
-
- Brotman DJ, Golden SH, Wittstein IS, The cardiovascular toll of stress. Lancet 370, 1089–1100 (2007). - PubMed
-
- Steptoe A, Kivimaki M, Stress and cardiovascular disease. Nature reviews. Cardiology 9, 360–370 (2012). - PubMed
-
- Kivimaki M, Pentti J, Ferrie JE, Batty GD, Nyberg ST, Jokela M, Virtanen M, Alfredsson L, Dragano N, Fransson EI, Goldberg M, Knutsson A, Koskenvuo M, Koskinen A, Kouvonen A, Luukkonen R, Oksanen T, Rugulies R, Siegrist J, Singh-Manoux A, Suominen S, Theorell T, Vaananen A, Vahtera J, Westerholm PJM, Westerlund H, Zins M, Strandberg T, Steptoe A, Deanfield J, consortium IP-W, Work stress and risk of death in men and women with and without cardiometabolic disease: a multicohort study. Lancet Diabetes Endocrinol, (2018). - PMC - PubMed
-
- Stewart RAH, Colquhoun DM, Marschner SL, Kirby AC, Simes J, Nestel PJ, Glozier N, O’Neil A, Oldenburg B, White HD, Tonkin AM, Investigators LS, Persistent psychological distress and mortality in patients with stable coronary artery disease. Heart 103, 1860–1866 (2017). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
