

RNA recognition motifs of disease-linked RNA-binding proteins contribute to amyloid formation
- PMID: 30992467
- PMCID: PMC6467989
- DOI: 10.1038/s41598-019-42367-8
RNA recognition motifs of disease-linked RNA-binding proteins contribute to amyloid formation
Abstract
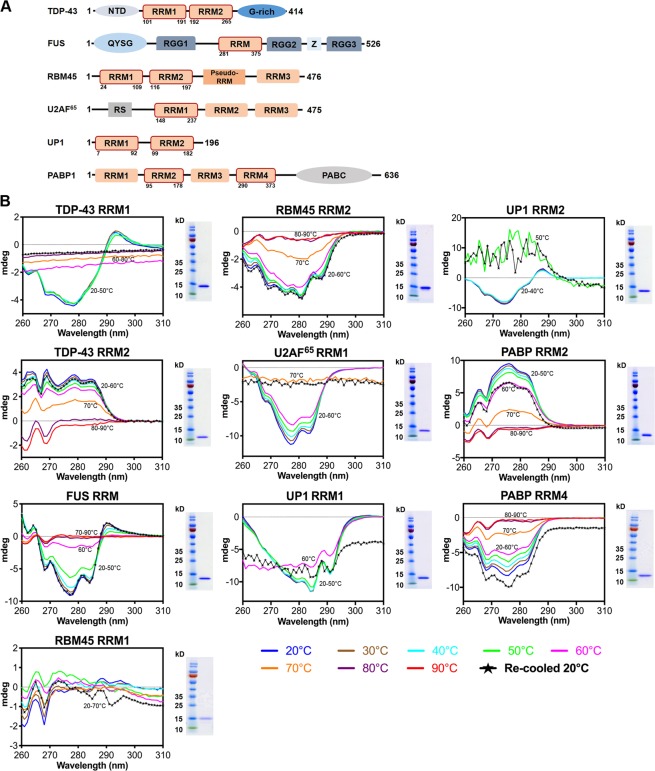
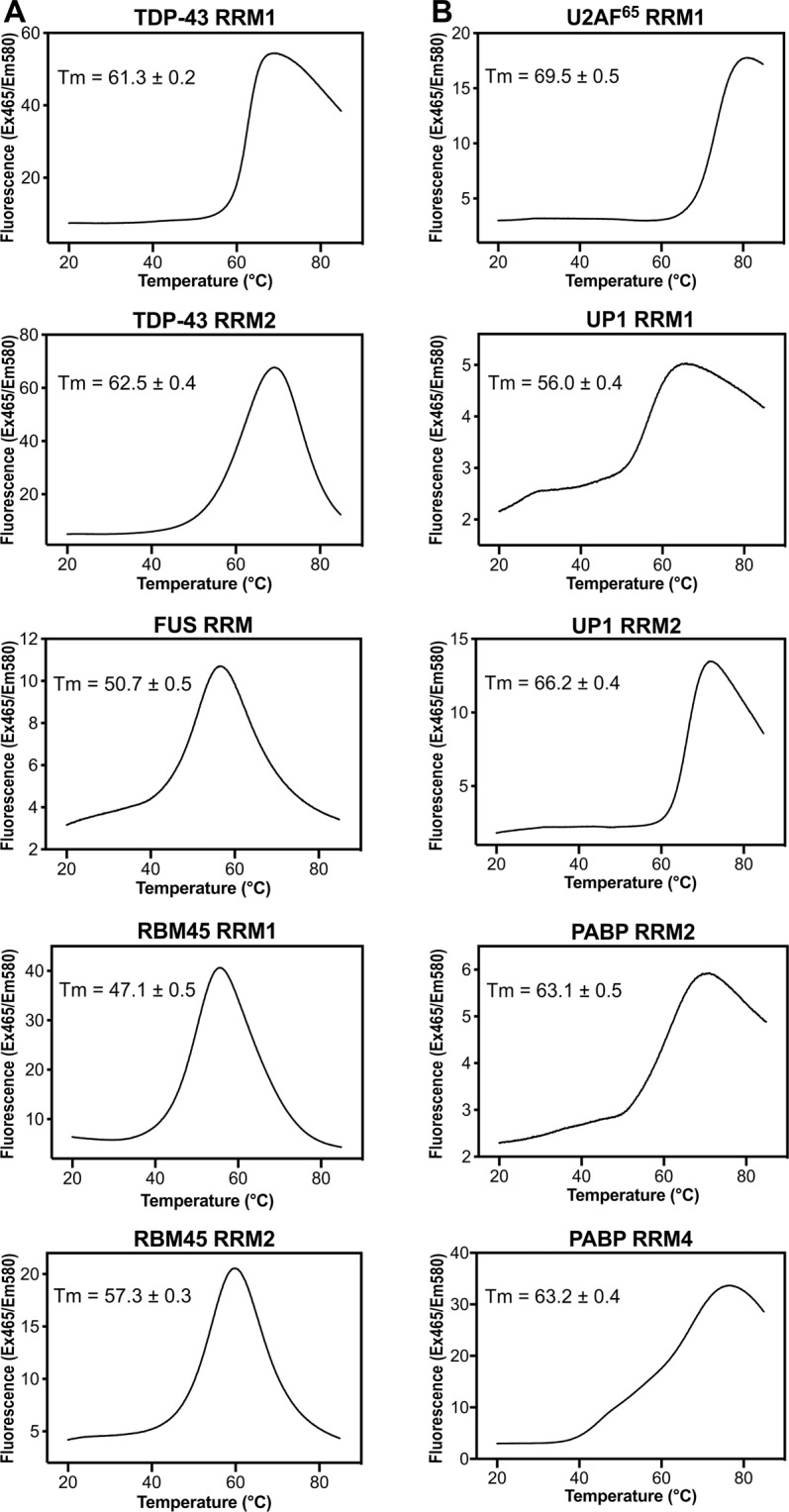
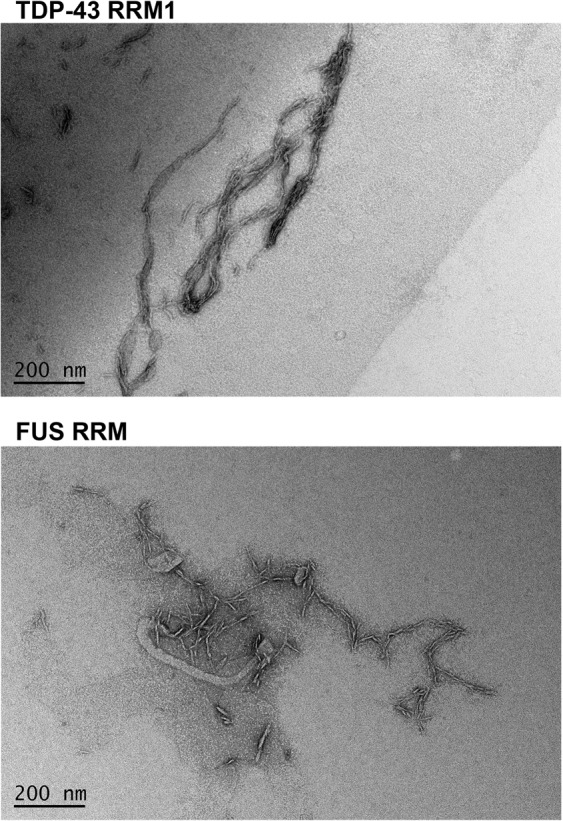
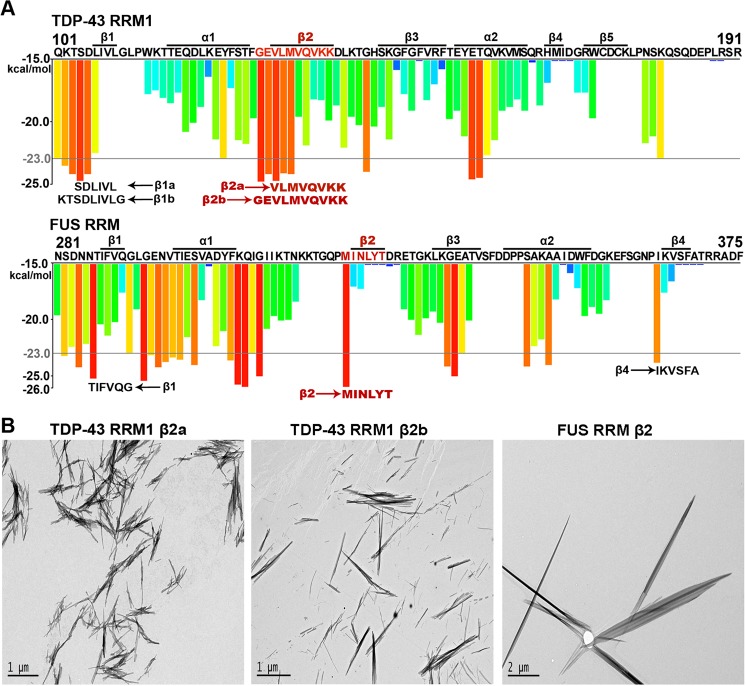
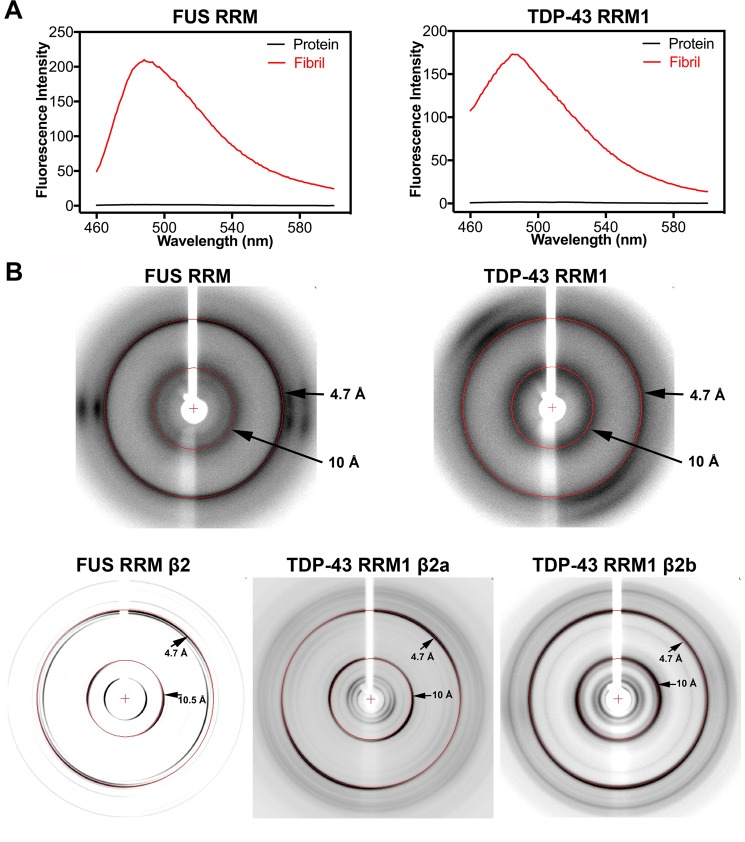
Aberrant expression, dysfunction and particularly aggregation of a group of RNA-binding proteins, including TDP-43, FUS and RBM45, are associated with neurological disorders. These three disease-linked RNA-binding proteins all contain at least one RNA recognition motif (RRM). However, it is not clear if these RRMs contribute to their aggregation-prone character. Here, we compare the biophysical and fibril formation properties of five RRMs from disease-linked RNA-binding proteins and five RRMs from non-disease-associated proteins to determine if disease-linked RRMs share specific features making them prone to self-assembly. We found that most of the disease-linked RRMs exhibit reversible thermal unfolding and refolding, and have a slightly lower average thermal melting point compared to that of normal RRMs. The full domain of TDP-43 RRM1 and FUS RRM, as well as the β-peptides from these two RRMs, could self-assemble into fibril-like aggregates which are amyloids of parallel β-sheets as verified by X-ray diffraction and FT-IR spectroscopy. Our results suggest that some disease-linked RRMs indeed play important roles in amyloid formation and shed light on why RNA-binding proteins with RRMs are frequently identified in the cellular inclusions of neurodegenerative diseases.
Conflict of interest statement
The authors declare no competing interests.
Figures

References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
