

Quality and quantity control of gene expression by nonsense-mediated mRNA decay
- PMID: 30992545
- PMCID: PMC6855384
- DOI: 10.1038/s41580-019-0126-2
Quality and quantity control of gene expression by nonsense-mediated mRNA decay
Erratum in
-
Publisher Correction: Quality and quantity control of gene expression by nonsense-mediated mRNA decay.Nat Rev Mol Cell Biol. 2019 Jun;20(6):384. doi: 10.1038/s41580-019-0138-y. Nat Rev Mol Cell Biol. 2019. PMID: 31028366
Abstract
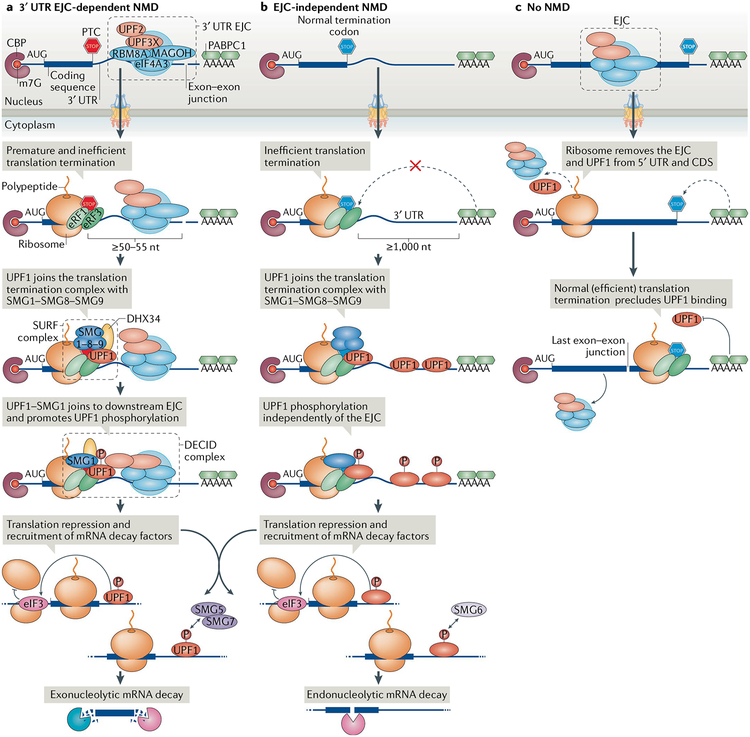
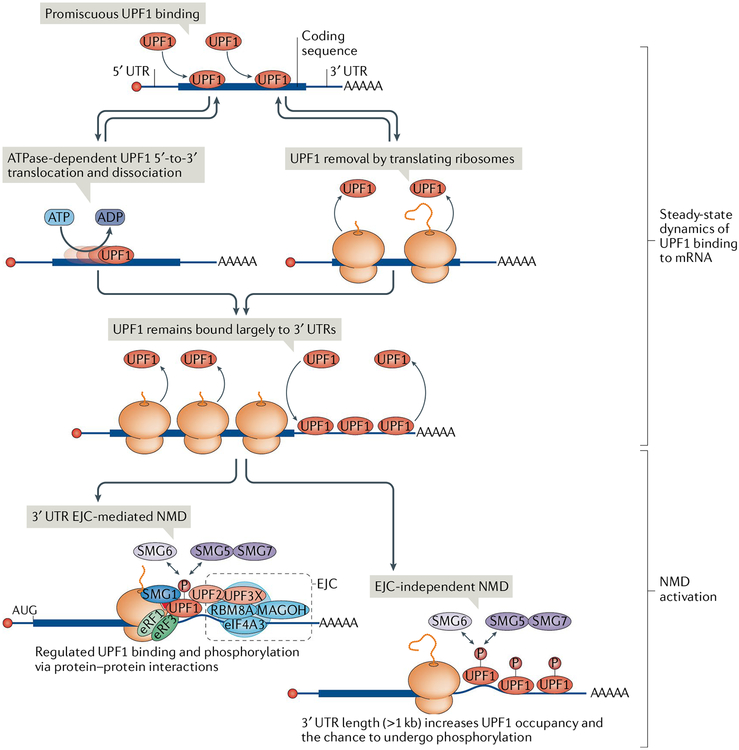
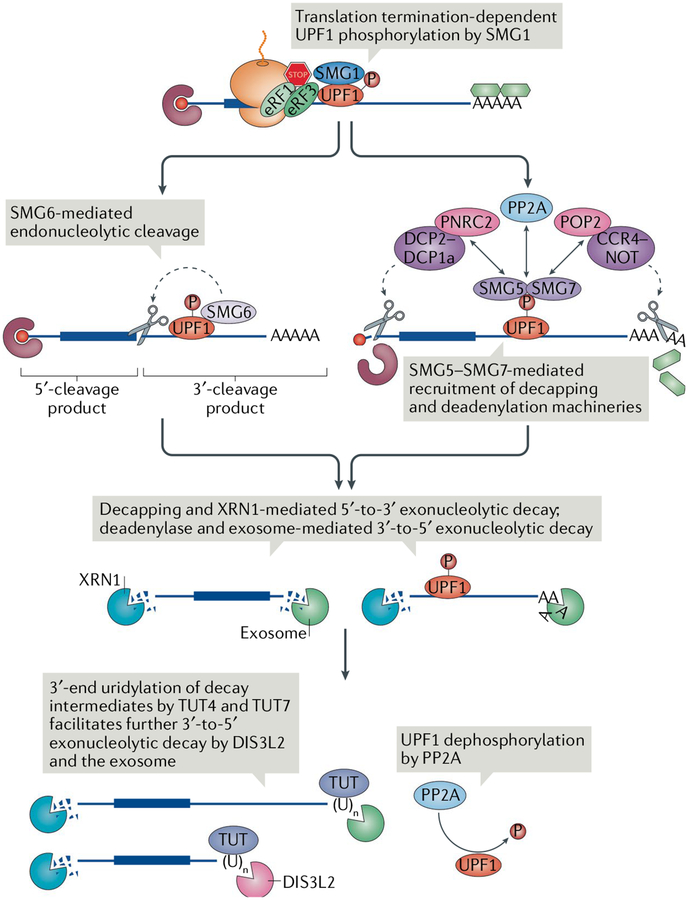
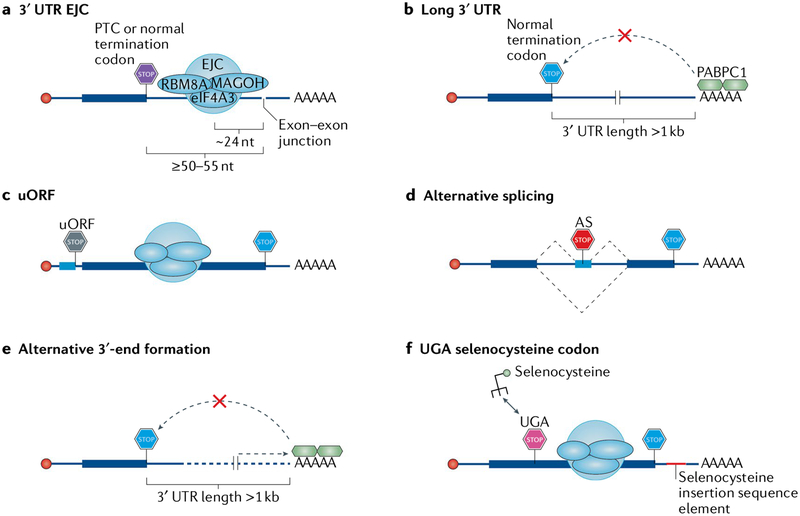
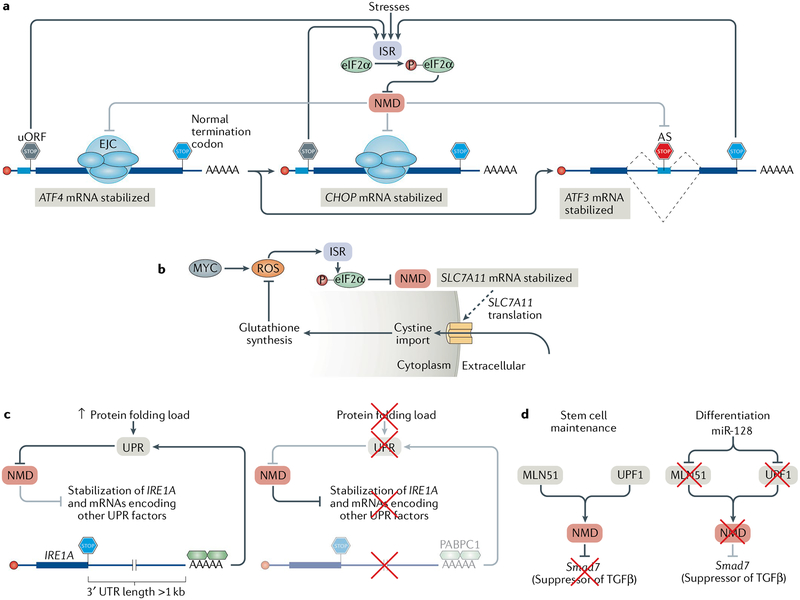
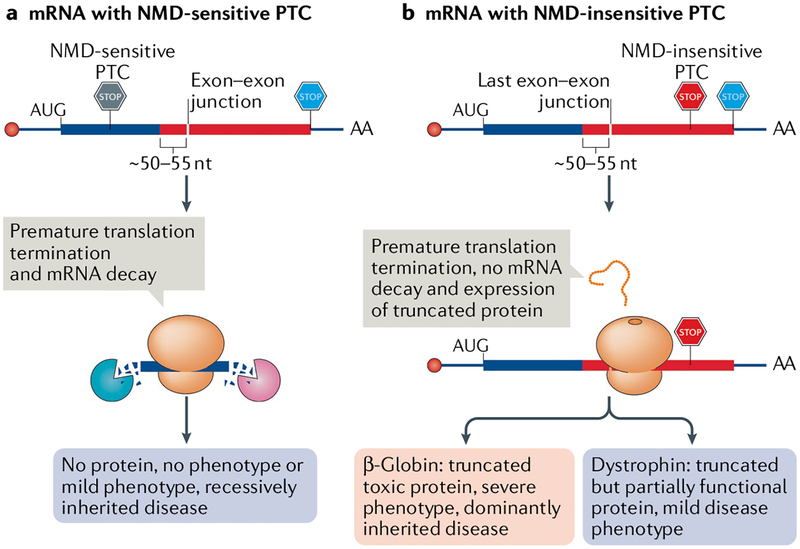
Nonsense-mediated mRNA decay (NMD) is one of the best characterized and most evolutionarily conserved cellular quality control mechanisms. Although NMD was first found to target one-third of mutated, disease-causing mRNAs, it is now known to also target ~10% of unmutated mammalian mRNAs to facilitate appropriate cellular responses - adaptation, differentiation or death - to environmental changes. Mutations in NMD genes in humans are associated with intellectual disability and cancer. In this Review, we discuss how NMD serves multiple purposes in human cells by degrading both mutated mRNAs to protect the integrity of the transcriptome and normal mRNAs to control the quantities of unmutated transcripts.
Conflict of interest statement
Competing interests
The authors declare no competing interests.
Figures
References
-
- Leeds P, Peltz SW, Jacobson A & Culbertson MR The product of the yeast UPF1 gene is required for rapid turnover of mRNAs containing a premature translational termination codon. Genes Dev 5, 2303–2314 (1991). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
