

In vivo assembly and trafficking of olfactory Ionotropic Receptors
- PMID: 30995910
- PMCID: PMC6472016
- DOI: 10.1186/s12915-019-0651-7
In vivo assembly and trafficking of olfactory Ionotropic Receptors
Abstract
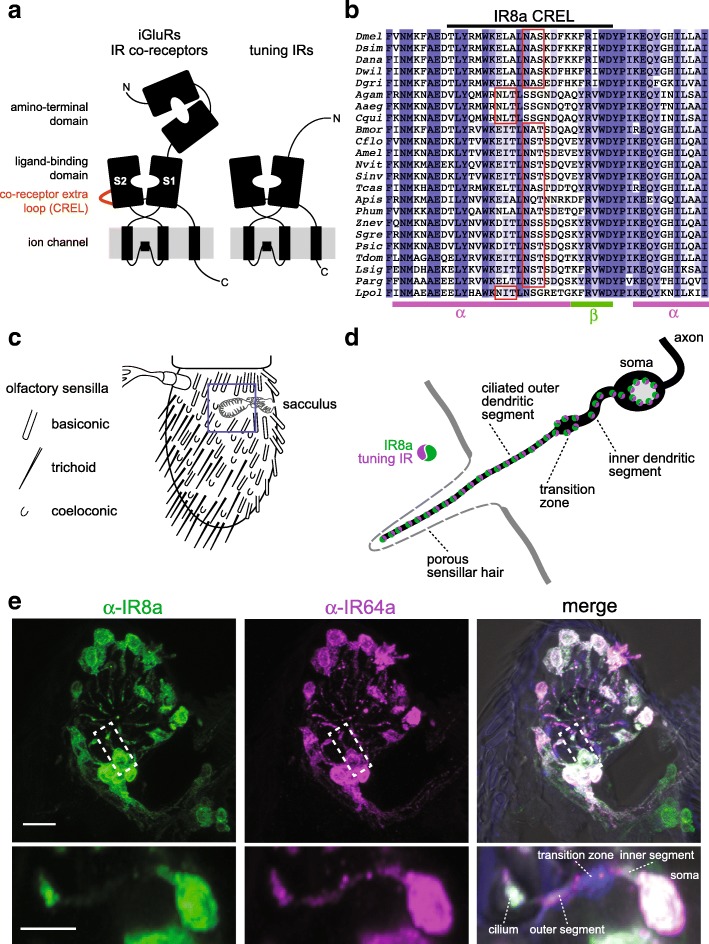
Background: Ionotropic receptors (IRs) are a large, divergent subfamily of ionotropic glutamate receptors (iGluRs) that are expressed in diverse peripheral sensory neurons and function in olfaction, taste, hygrosensation and thermosensation. Analogous to the cell biological properties of their synaptic iGluR ancestors, IRs are thought to form heteromeric complexes that localise to the ciliated dendrites of sensory neurons. IR complexes are composed of selectively expressed 'tuning' receptors and one of two broadly expressed co-receptors (IR8a or IR25a). While the extracellular ligand-binding domain (LBD) of tuning IRs is likely to define the stimulus specificity of the complex, the role of this domain in co-receptors is unclear.
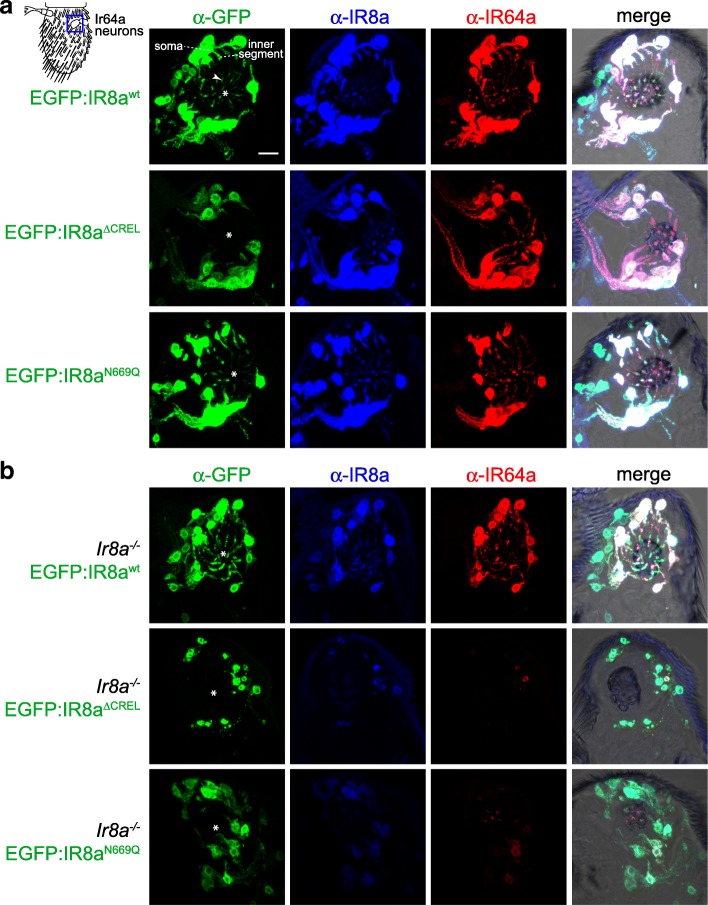
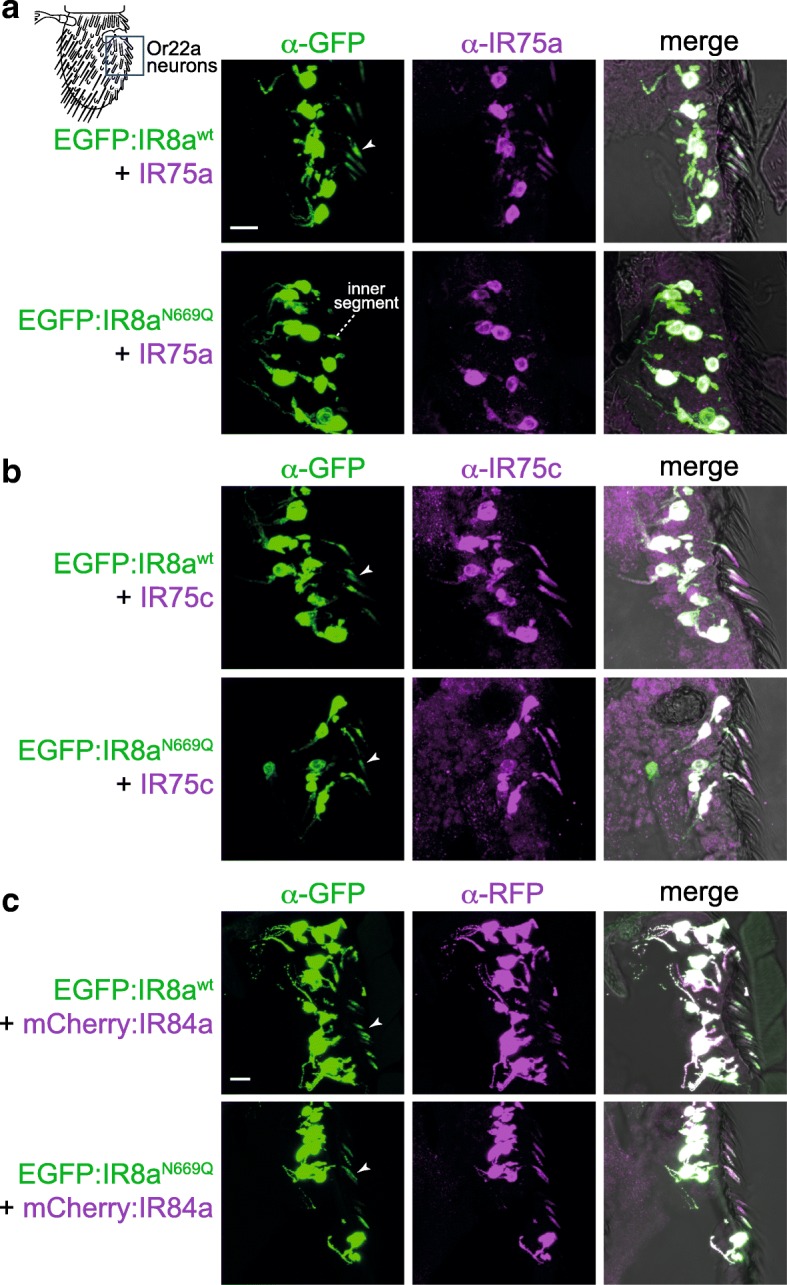
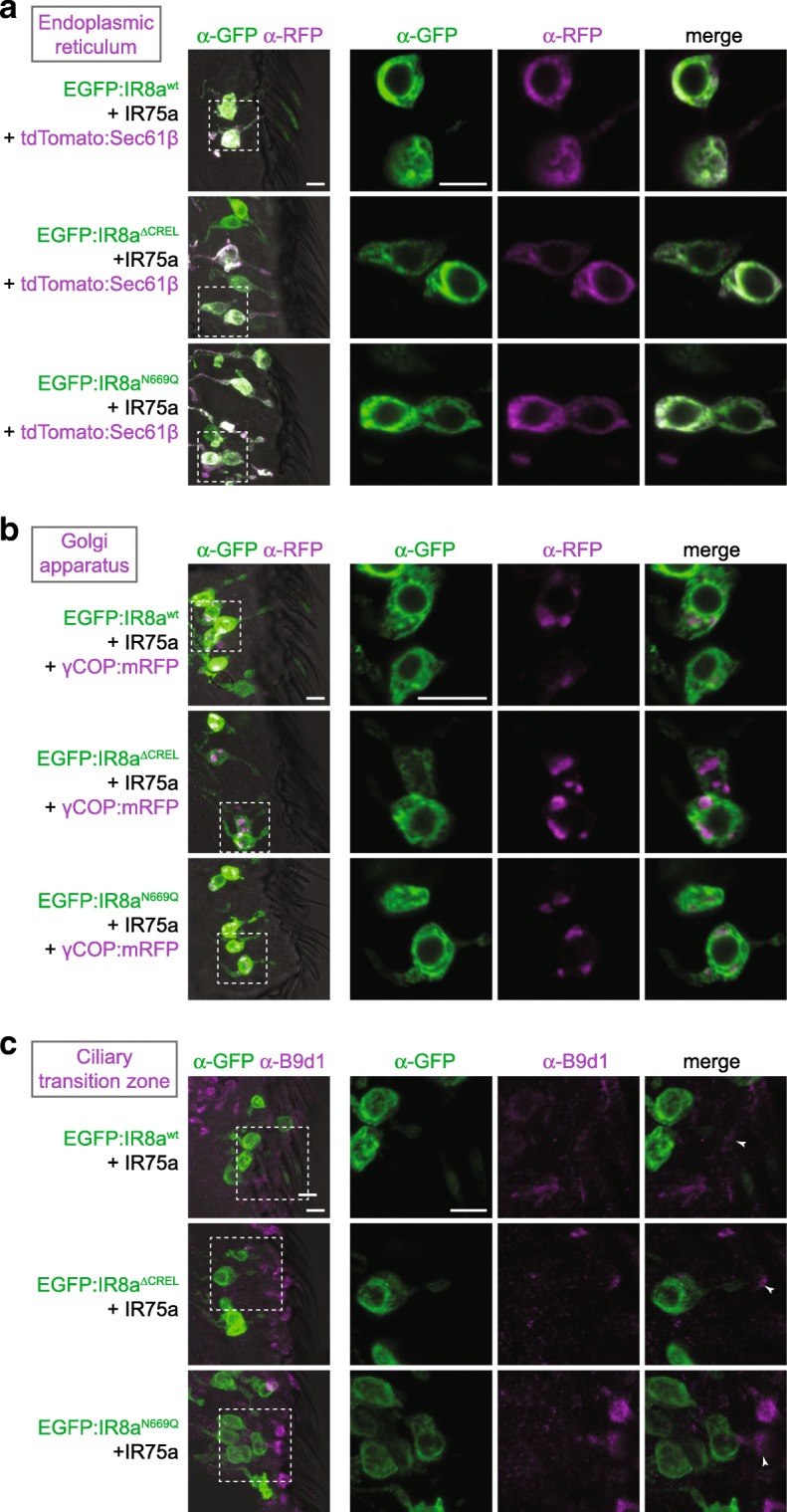
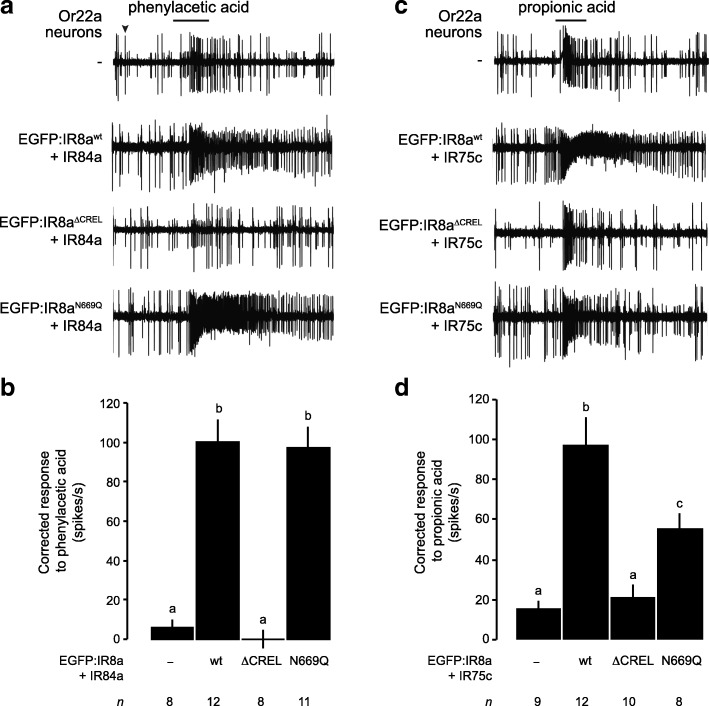
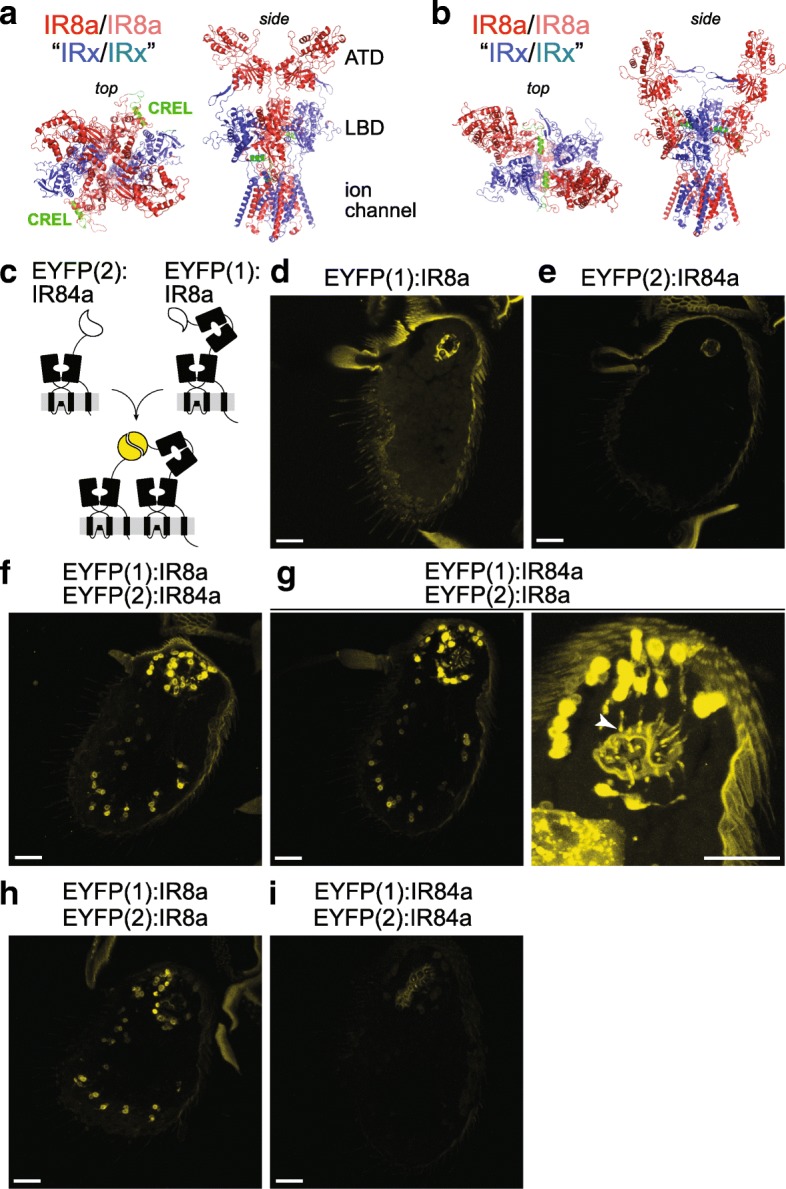
Results: We identify a sequence in the co-receptor LBD, the 'co-receptor extra loop' (CREL), which is conserved across IR8a and IR25a orthologues but not present in either tuning IRs or iGluRs. The CREL contains a single predicted N-glycosylation site, which we show bears a sugar modification in recombinantly expressed IR8a. Using the Drosophila olfactory system as an in vivo model, we find that a transgenically encoded IR8a mutant in which the CREL cannot be N-glycosylated is impaired in localisation to cilia in some, though not all, populations of sensory neurons expressing different tuning IRs. This defect can be complemented by the presence of endogenous wild-type IR8a, indicating that IR complexes contain at least two IR8a subunits and that this post-translational modification is dispensable for protein folding or complex assembly. Analysis of the subcellular distribution of the mutant protein suggests that its absence from sensory cilia is due to a failure in exit from the endoplasmic reticulum. Protein modelling and in vivo analysis of tuning IR and co-receptor subunit interactions by a fluorescent protein fragment complementation assay reveal that the CREL N-glycosylation site is likely to be located on the external face of a heterotetrameric IR complex.
Conclusions: Our data reveal an important role for the IR co-receptor LBD in control of intracellular transport, provide novel insights into the stoichiometry and assembly of IR complexes and uncover an unexpected heterogeneity in the trafficking regulation of this sensory receptor family.
Conflict of interest statement
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
References
-
- Gereau RW, Swanson GT. The glutamate receptors. Totowa: Humana Press; 2008.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
