

Human antibody response to Zika targets type-specific quaternary structure epitopes
- PMID: 30996133
- PMCID: PMC6538335
- DOI: 10.1172/jci.insight.124588
Human antibody response to Zika targets type-specific quaternary structure epitopes
Abstract
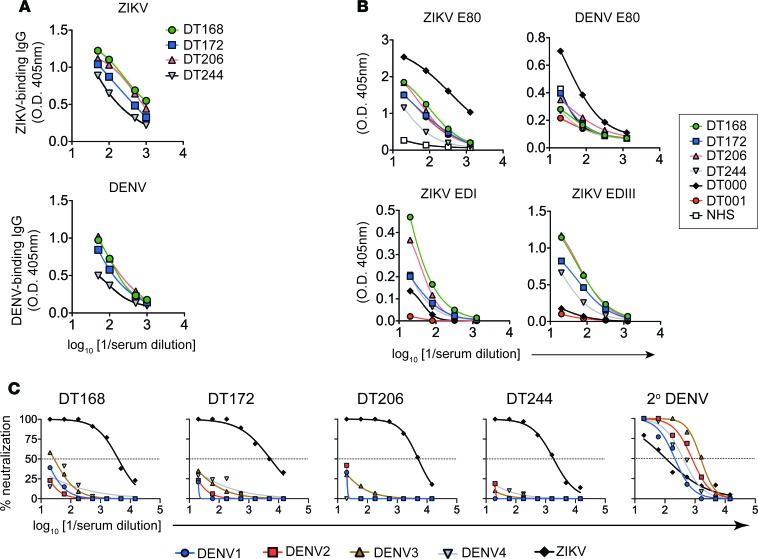
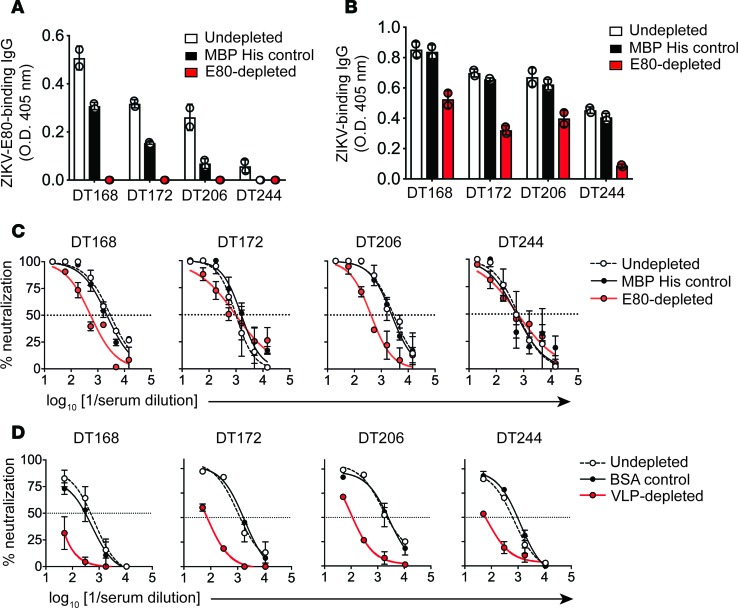
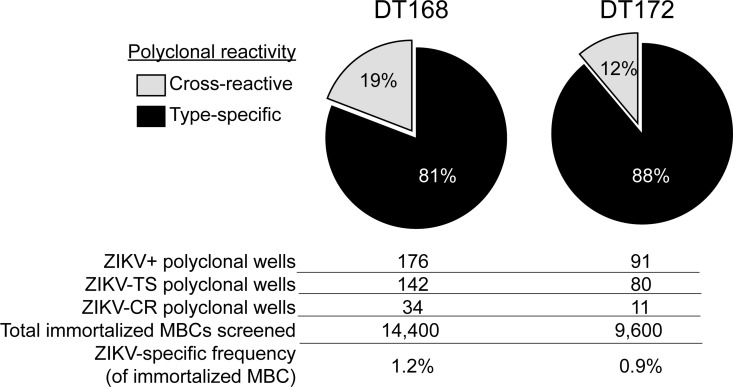
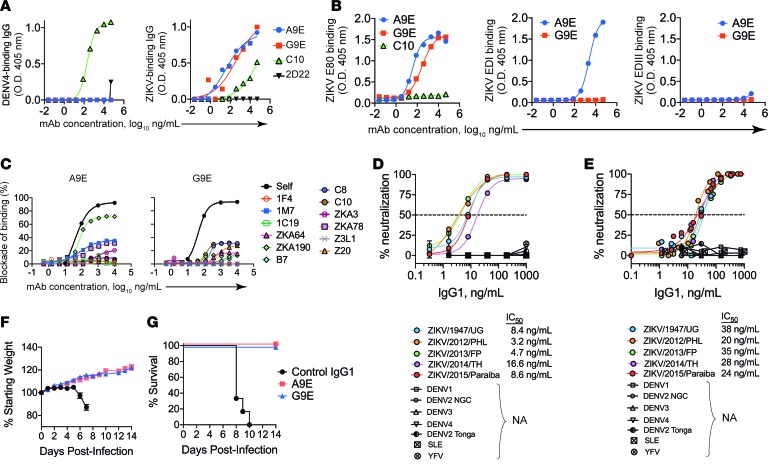
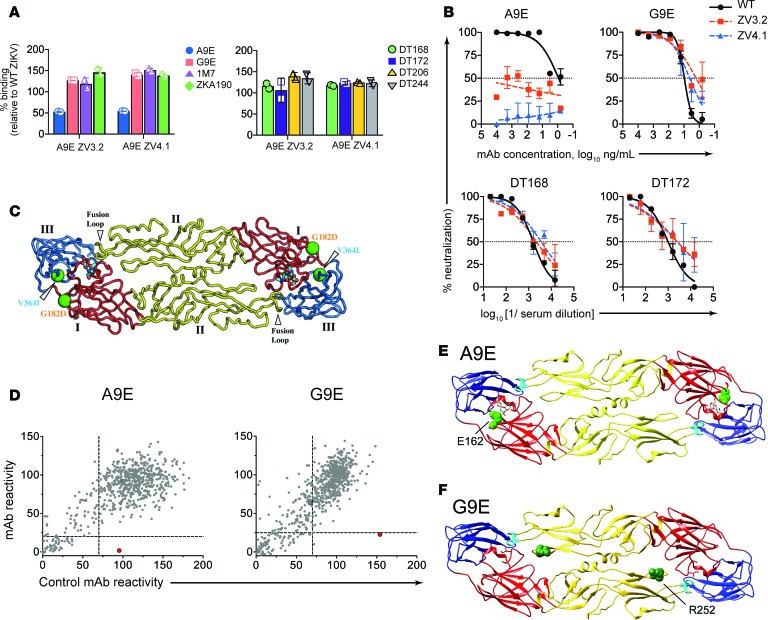
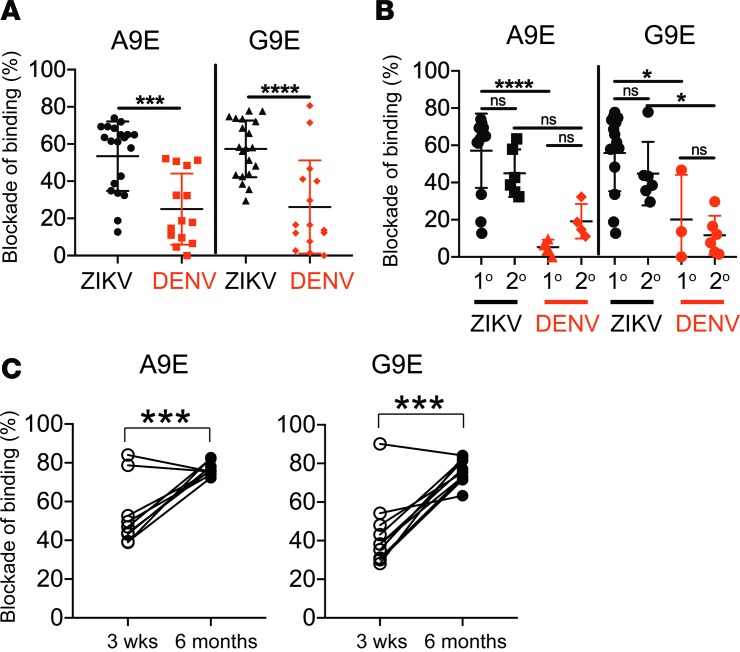
The recent Zika virus (ZIKV) epidemic in the Americas has revealed rare but serious manifestations of infection. ZIKV has emerged in regions endemic for dengue virus (DENV), a closely related mosquito-borne flavivirus. Cross-reactive antibodies confound studies of ZIKV epidemiology and pathogenesis. The immune responses to ZIKV may be different in people, depending on their DENV immune status. Here, we focus on the human B cell and antibody response to ZIKV as a primary flavivirus infection to define the properties of neutralizing and protective antibodies generated in the absence of preexisting immunity to DENV. The plasma antibody and memory B cell response is highly ZIKV type-specific, and ZIKV-neutralizing antibodies mainly target quaternary structure epitopes on the viral envelope. To map viral epitopes targeted by protective antibodies, we isolated 2 type-specific monoclonal antibodies (mAbs) from a ZIKV case. Both mAbs were strongly neutralizing in vitro and protective in vivo. The mAbs recognize distinct epitopes centered on domains I and II of the envelope protein. We also demonstrate that the epitopes of these mAbs define antigenic regions commonly targeted by plasma antibodies in individuals from endemic and nonendemic regions who have recovered from ZIKV infections.
Keywords: Adaptive immunity; B cells; Immunoglobulins; Immunology; Virology.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
- P20 RR021905/RR/NCRR NIH HHS/United States
- T32 AI007151/AI/NIAID NIH HHS/United States
- P30 CA016086/CA/NCI NIH HHS/United States
- R01 HD094009/HD/NICHD NIH HHS/United States
- R21 AI134073/AI/NIAID NIH HHS/United States
- P20 GM125498/GM/NIGMS NIH HHS/United States
- T32 AI055402/AI/NIAID NIH HHS/United States
- P30 GM118228/GM/NIGMS NIH HHS/United States
- HHSN272201400058C/AI/NIAID NIH HHS/United States
- R21 AI129532/AI/NIAID NIH HHS/United States
- S10 OD018175/OD/NIH HHS/United States
- R01 AI107731/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
