

CARD9+ microglia promote antifungal immunity via IL-1β- and CXCL1-mediated neutrophil recruitment
- PMID: 30996332
- PMCID: PMC6494474
- DOI: 10.1038/s41590-019-0377-2
CARD9+ microglia promote antifungal immunity via IL-1β- and CXCL1-mediated neutrophil recruitment
Abstract
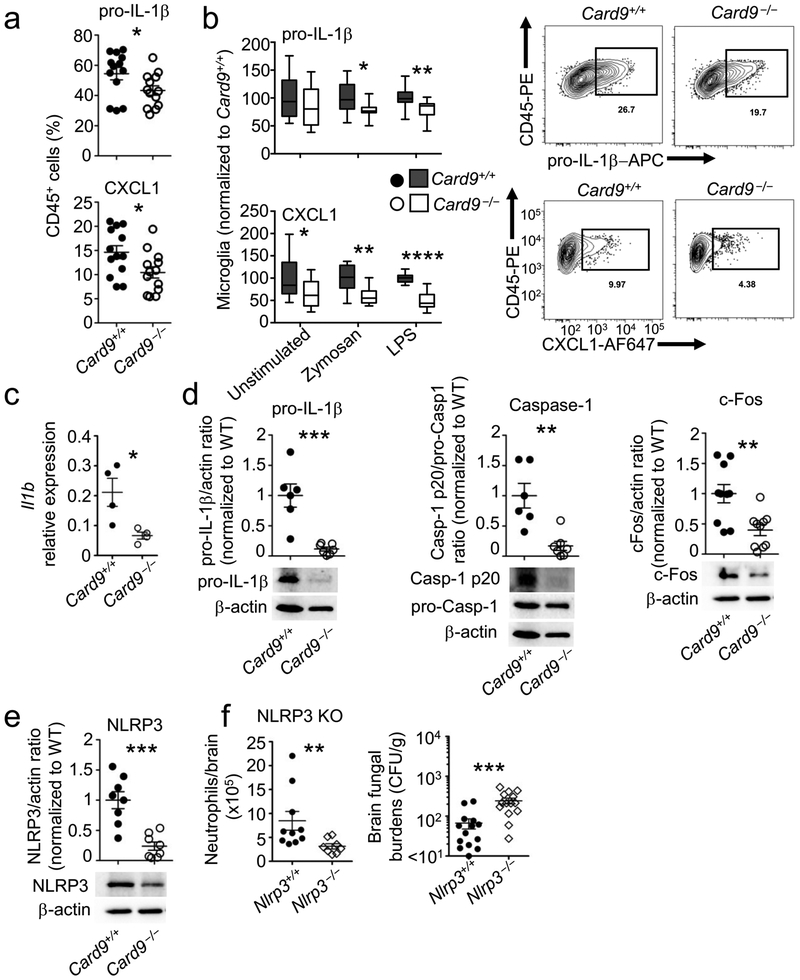
The C-type lectin receptor-Syk (spleen tyrosine kinase) adaptor CARD9 facilitates protective antifungal immunity within the central nervous system (CNS), as human deficiency in CARD9 causes susceptibility to fungus-specific, CNS-targeted infection. CARD9 promotes the recruitment of neutrophils to the fungus-infected CNS, which mediates fungal clearance. In the present study we investigated host and pathogen factors that promote protective neutrophil recruitment during invasion of the CNS by Candida albicans. The cytokine IL-1β served an essential function in CNS antifungal immunity by driving production of the chemokine CXCL1, which recruited neutrophils expressing the chemokine receptor CXCR2. Neutrophil-recruiting production of IL-1β and CXCL1 was induced in microglia by the fungus-secreted toxin Candidalysin, in a manner dependent on the kinase p38 and the transcription factor c-Fos. Notably, microglia relied on CARD9 for production of IL-1β, via both transcriptional regulation of Il1b and inflammasome activation, and of CXCL1 in the fungus-infected CNS. Microglia-specific Card9 deletion impaired the production of IL-1β and CXCL1 and neutrophil recruitment, and increased fungal proliferation in the CNS. Thus, an intricate network of host-pathogen interactions promotes antifungal immunity in the CNS; this is impaired in human deficiency in CARD9, which leads to fungal disease of the CNS.
Figures







Comment in
-
Microglia and Neutrophils to the Rescue.Trends Immunol. 2019 Jul;40(7):555-556. doi: 10.1016/j.it.2019.05.005. Epub 2019 May 24. Trends Immunol. 2019. PMID: 31133466 Free PMC article.
References
-
- Lionakis MS & Levitz SM Host control of fungal infections: lessons from basic studies and human cohorts. Ann Rev Immunol 36, 157–191 (2018). - PubMed
Methods-only References
-
- Ruland J, Duncan GS, Wakeham A, Mak TW Differential requirement for Malt1 in T and B cell antigen receptor signaling. Immunity 19(5), 749–58 (2003) - PubMed
-
- Tay TL, Mai D, Dautzenberg J, Fernandez-Klett F, Lin G et al. A new fate mapping system reveals context-dependent random or clonal expansion of microglia. Nat Neurosci 20, 793–803 (2017) - PubMed
-
- Goldmann T, Wieghofer P, Muller PF, Wolf Y, Varol D et al. A new type of microglia gene targeting shows TAK1 to be pivotal in CNS autoimmune inflammation. Nat Neurosci 16, 1618–1626 (2013) - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 DE026600/DE/NIDCR NIH HHS/United States
- R37 AI093808/AI/NIAID NIH HHS/United States
- R01 CA161373/CA/NCI NIH HHS/United States
- P30 CA008748/CA/NCI NIH HHS/United States
- MR/M011372/1/MRC_/Medical Research Council/United Kingdom
- R01 093808/NH/NIH HHS/United States
- R01CA161373/NH/NIH HHS/United States
- R01 DE017088/DE/NIDCR NIH HHS/United States
- 102705/Z/13/Z/WT_/Wellcome Trust/United Kingdom
- MR/N006364/2/MRC_/Medical Research Council/United Kingdom
- ZIA AI001175/ImNIH/Intramural NIH HHS/United States
- R01 DK110352/DK/NIDDK NIH HHS/United States
- R01 AI124566/AI/NIAID NIH HHS/United States
- R01AI124566/NH/NIH HHS/United States
- MR/N006364/1/MRC_/Medical Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
