

Pitaya HpWRKY3 Is Associated with Fruit Sugar Accumulation by Transcriptionally Modulating Sucrose Metabolic Genes HpINV2 and HpSuSy1
- PMID: 30999552
- PMCID: PMC6514986
- DOI: 10.3390/ijms20081890
Pitaya HpWRKY3 Is Associated with Fruit Sugar Accumulation by Transcriptionally Modulating Sucrose Metabolic Genes HpINV2 and HpSuSy1
Abstract
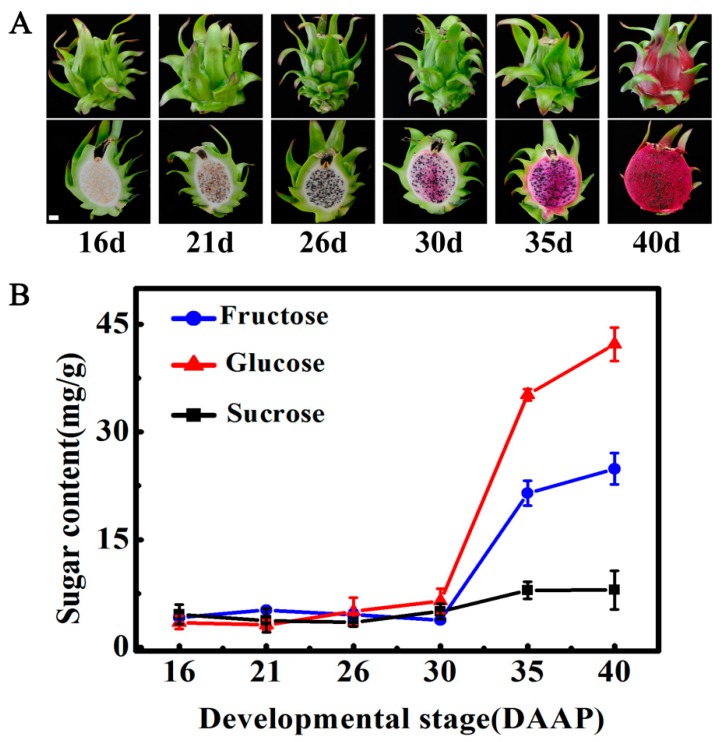
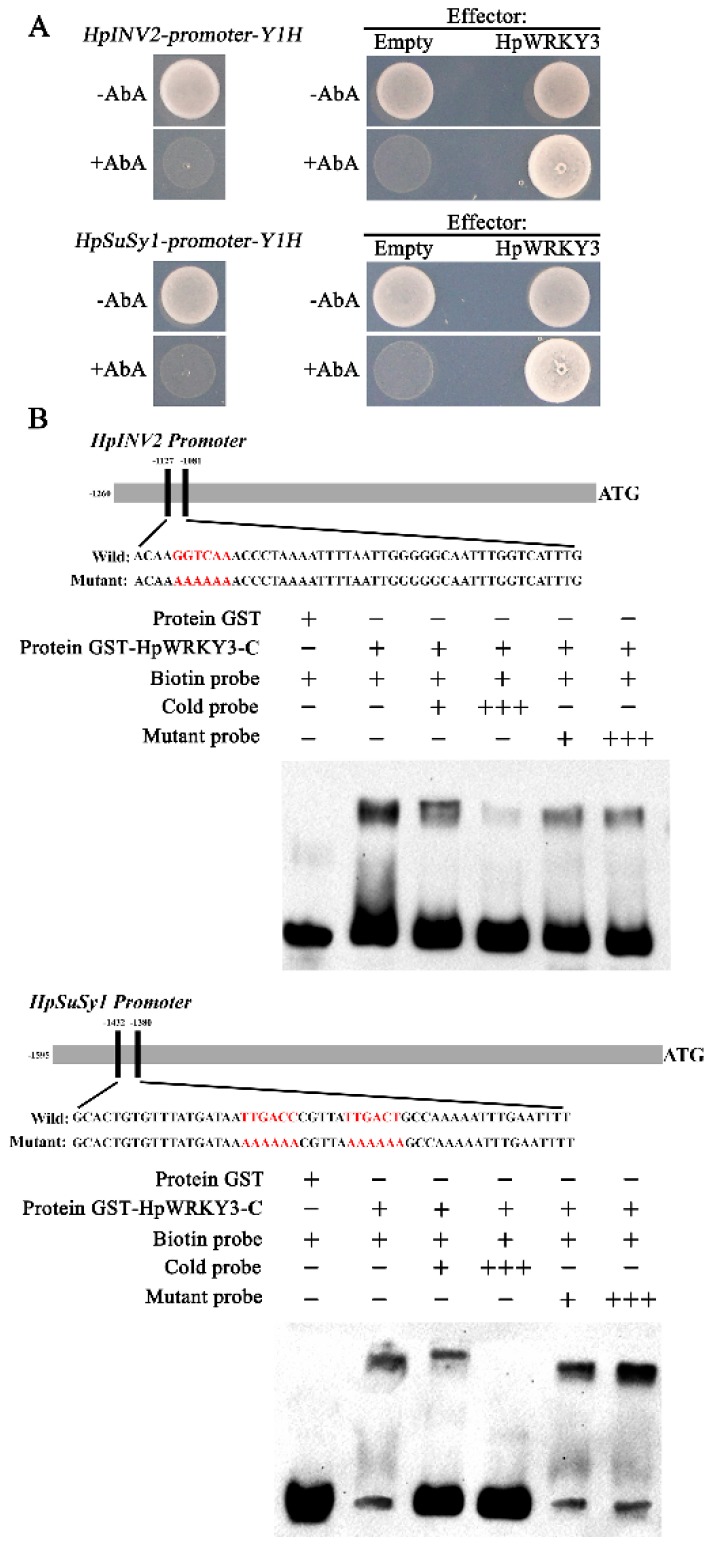
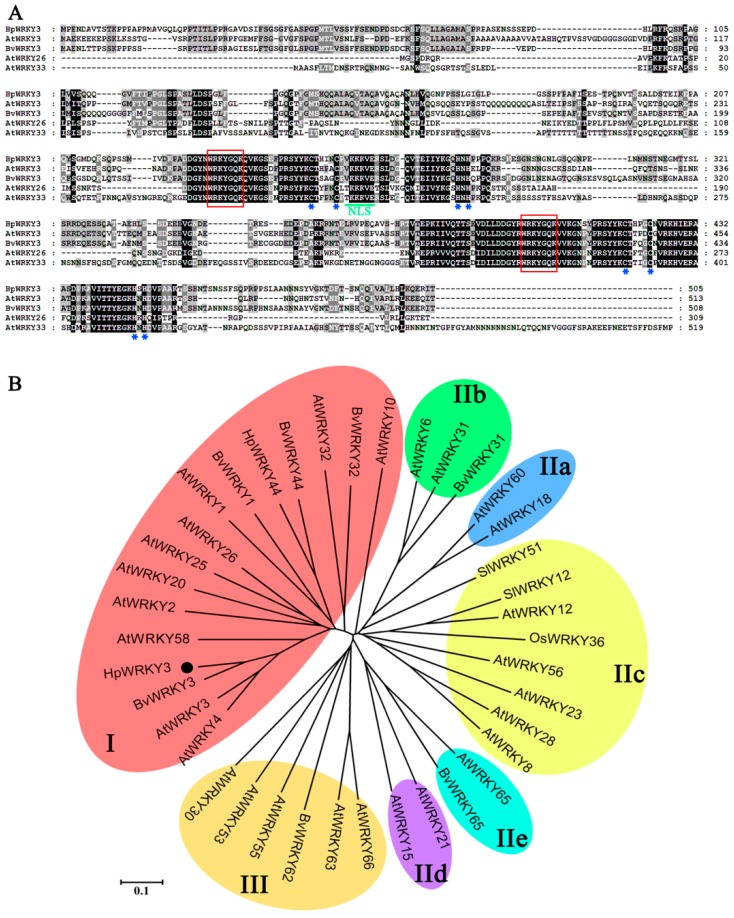
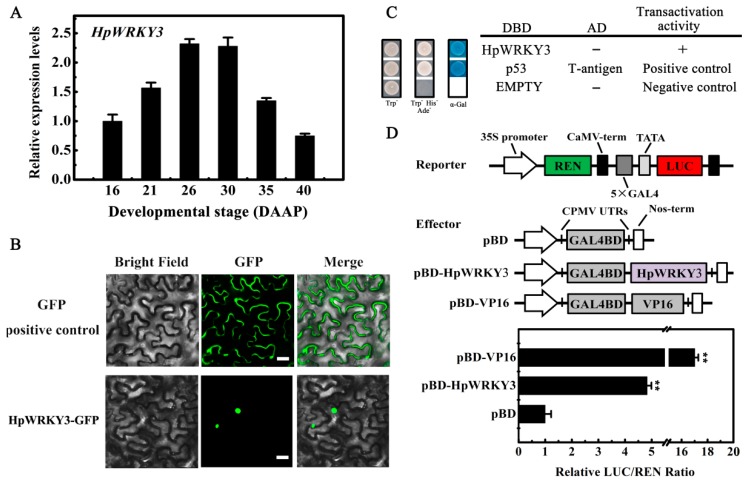
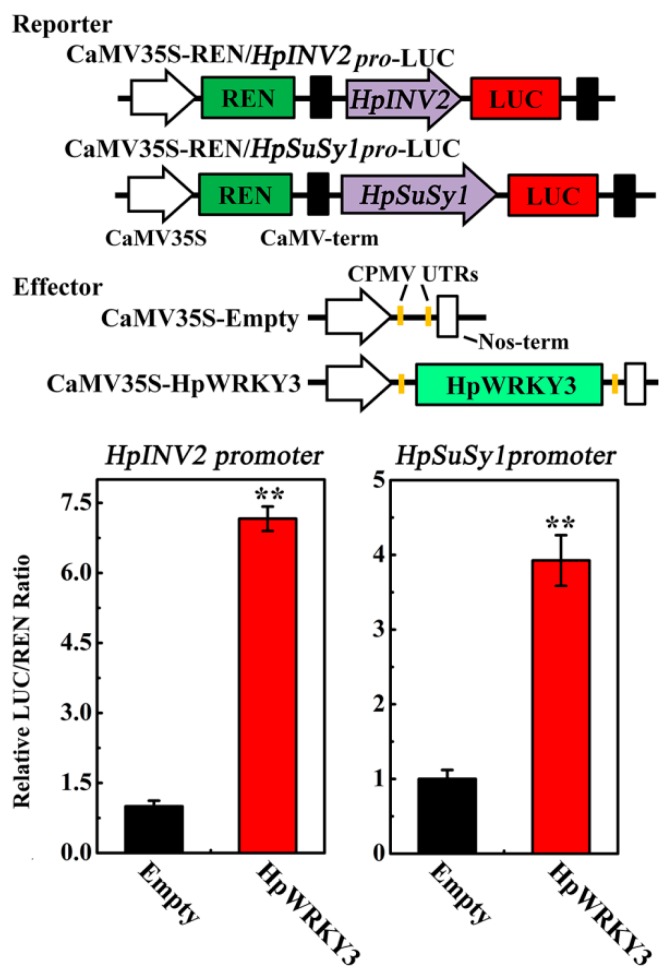
Sugar level is an important determinant of fruit taste and consumer preferences. However, upstream regulators that control sugar accumulation during fruit maturation are poorly understood. In the present work, we found that glucose is the main sugar in mature pitaya (Hylocereus) fruit, followed by fructose and sucrose. Expression levels of two sucrose-hydrolyzing enzyme genes HpINV2 and HpSuSy1 obviously increased during fruit maturation, which were correlated well with the elevated accumulation of glucose and fructose. A WRKY transcription factor HpWRKY3 was further identified as the putative binding protein of the HpINV2 and HpSuSy1 promoters by yeast one-hybrid and gel mobility shift assays. HpWRKY3 was localized exclusively in the nucleus and possessed trans-activation ability. HpWRKY3 exhibited the similar expression pattern with HpINV2 and HpSuSy1. Finally, transient expression assays in tobacco leaves showed that HpWRKY3 activated the expressions of HpINV2 and HpSuSy1. Taken together, we propose that HpWRKY3 is associated with pitaya fruit sugar accumulation by activating the transcriptions of sucrose metabolic genes. Our findings thus shed light on the transcriptional mechanism that regulates the sugar accumulation during pitaya fruit quality formation.
Keywords: fruit maturation; fruit quality; sucrose-hydrolyzing enzyme genes; transcriptional activation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- García-Cruz L., Dueñas M., Santos-Buelgas C., Valle-Guadarrama S., Salinas-Moreno Y. Betalains and phenolic compounds profiling and antioxidant capacity of pitaya (Stenocereus spp.) fruit from two species (S. Pruinosus and S. stellatus) Food Chem. 2017;234:111–118. doi: 10.1016/j.foodchem.2017.04.174. - DOI - PubMed
-
- Menezes Cordeiro M.H., Da Silva J.M., Mizobutsi G.P., Mizobutsi E.H., Da Mota W.F. Physical, chemical and nutritional characterization of pink pitaya of red pulp. Rev. Bras. Frutic. 2015;37:20–26.
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
