

Complex signal processing in synthetic gene circuits using cooperative regulatory assemblies
- PMID: 31000590
- PMCID: PMC6650298
- DOI: 10.1126/science.aau8287
Complex signal processing in synthetic gene circuits using cooperative regulatory assemblies
Abstract
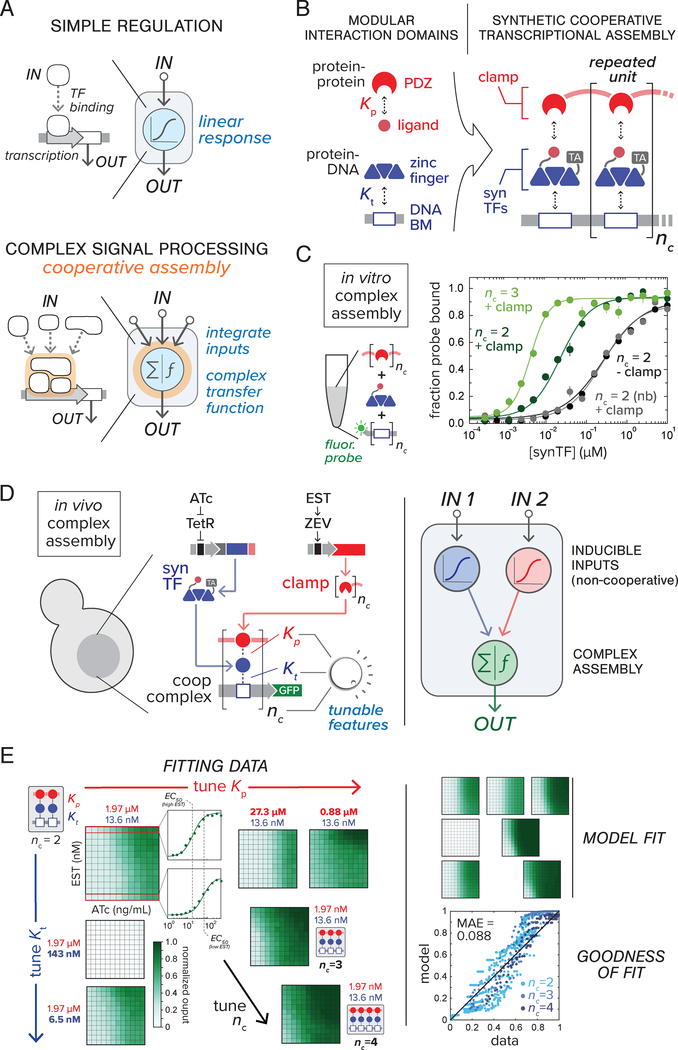
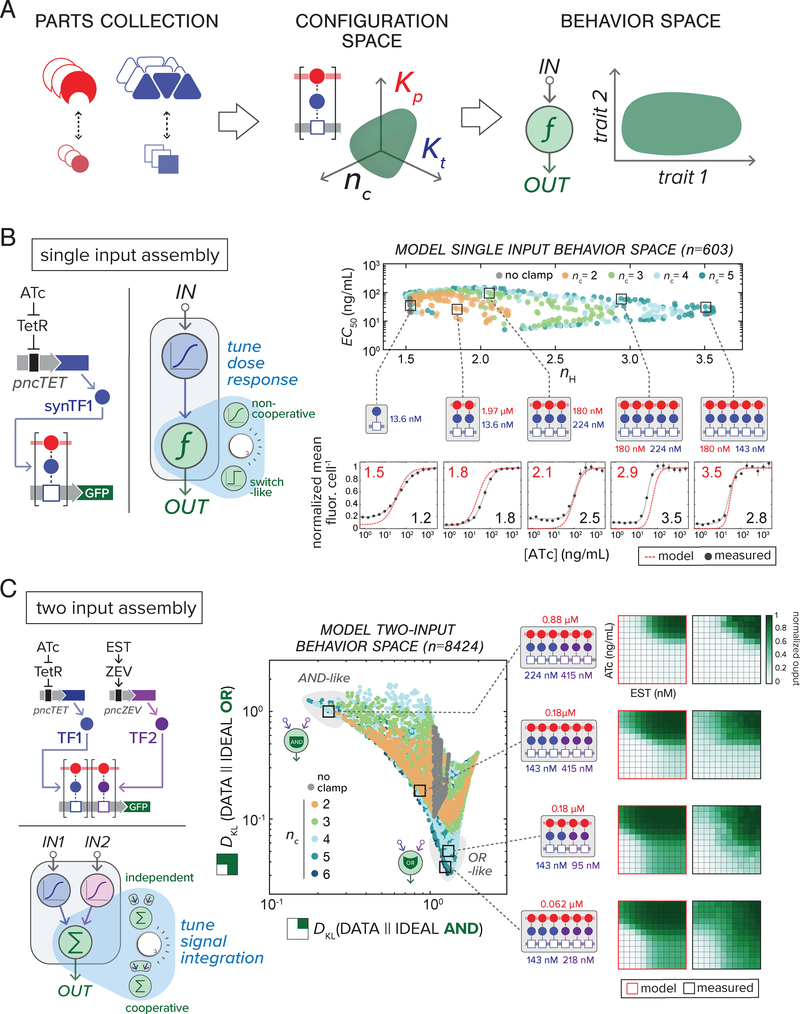
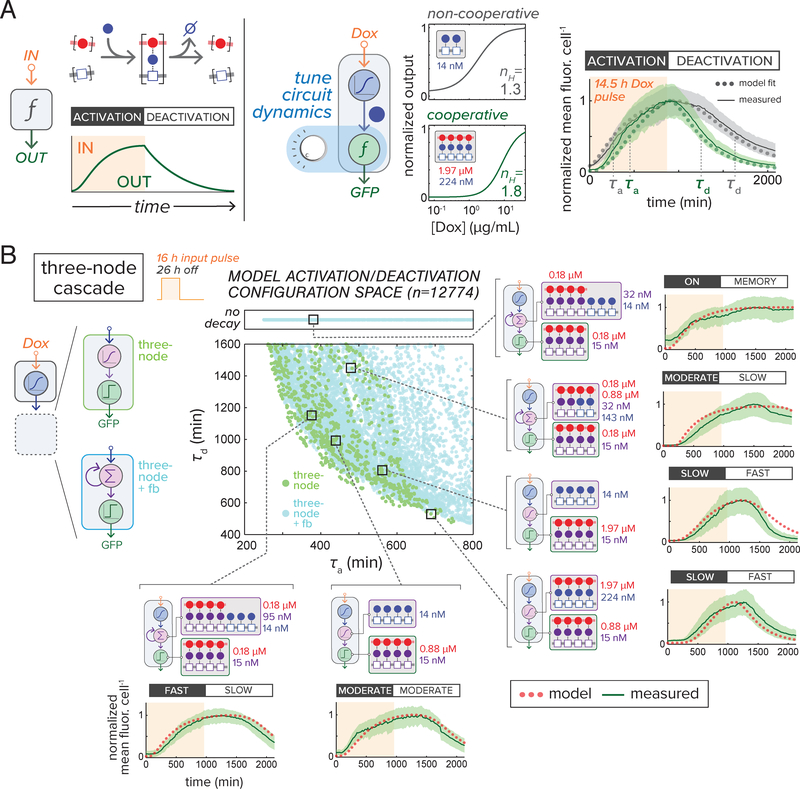
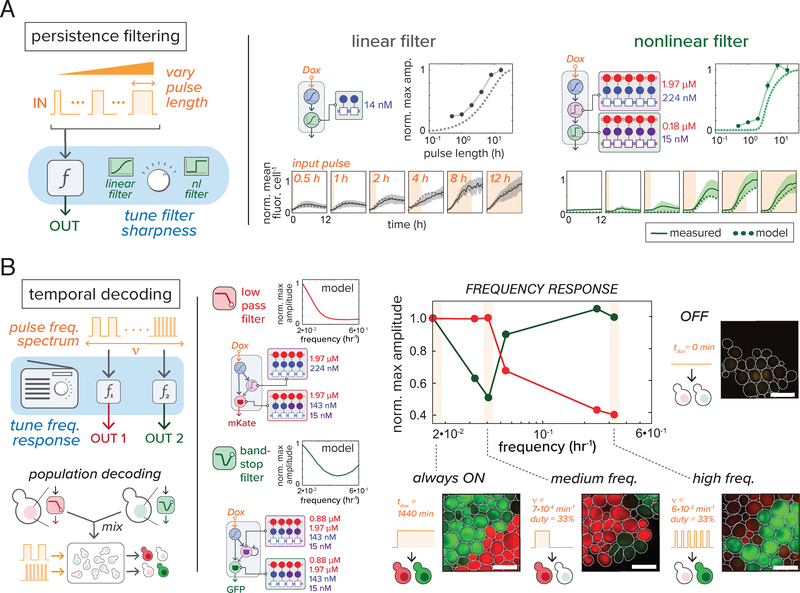
Eukaryotic genes are regulated by multivalent transcription factor complexes. Through cooperative self-assembly, these complexes perform nonlinear regulatory operations involved in cellular decision-making and signal processing. In this study, we apply this design principle to synthetic networks, testing whether engineered cooperative assemblies can program nonlinear gene circuit behavior in yeast. Using a model-guided approach, we show that specifying the strength and number of assembly subunits enables predictive tuning between linear and nonlinear regulatory responses for single- and multi-input circuits. We demonstrate that assemblies can be adjusted to control circuit dynamics. We harness this capability to engineer circuits that perform dynamic filtering, enabling frequency-dependent decoding in cell populations. Programmable cooperative assembly provides a versatile way to tune the nonlinearity of network connections, markedly expanding the engineerable behaviors available to synthetic circuits.
Copyright © 2019 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Figures
Comment in
-
Synthetic transcriptional synergy.Science. 2019 May 10;364(6440):531-532. doi: 10.1126/science.aax4556. Science. 2019. PMID: 31073055 No abstract available.
References
-
- Hill AV, The possible effects of the aggregation of the molecules of haemoglobin on its dissociation curves. J Physiol. 40, iv–vii (1910).
-
- Whitty A, Cooperativity and biological complexity. Nat Chem Biol. 4, 435–9 (2008). - PubMed
-
- Ptashne M, How eukaryotic transcriptional activators work. Nature. 335, 683–9 (1988). - PubMed
-
- Bhalla US and Iyengar R, Emergent properties of networks of biological signaling pathways. Science. 283, 381–7 (1999). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
