

A Mobile Auxin Signal Connects Temperature Sensing in Cotyledons with Growth Responses in Hypocotyls
- PMID: 31000634
- PMCID: PMC6548272
- DOI: 10.1104/pp.18.01377
A Mobile Auxin Signal Connects Temperature Sensing in Cotyledons with Growth Responses in Hypocotyls
Abstract
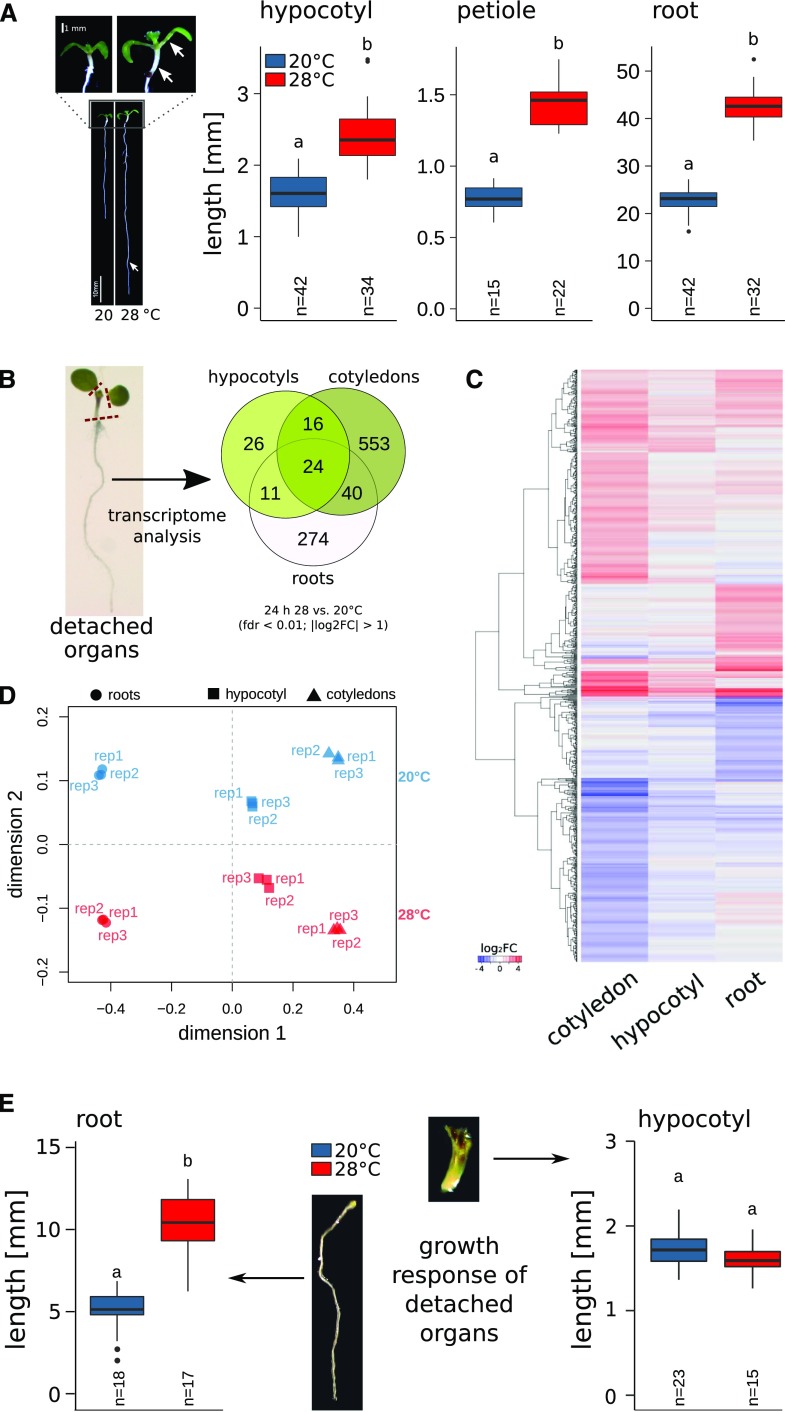
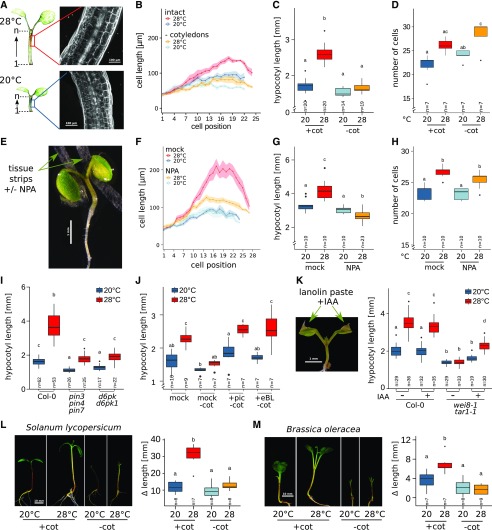
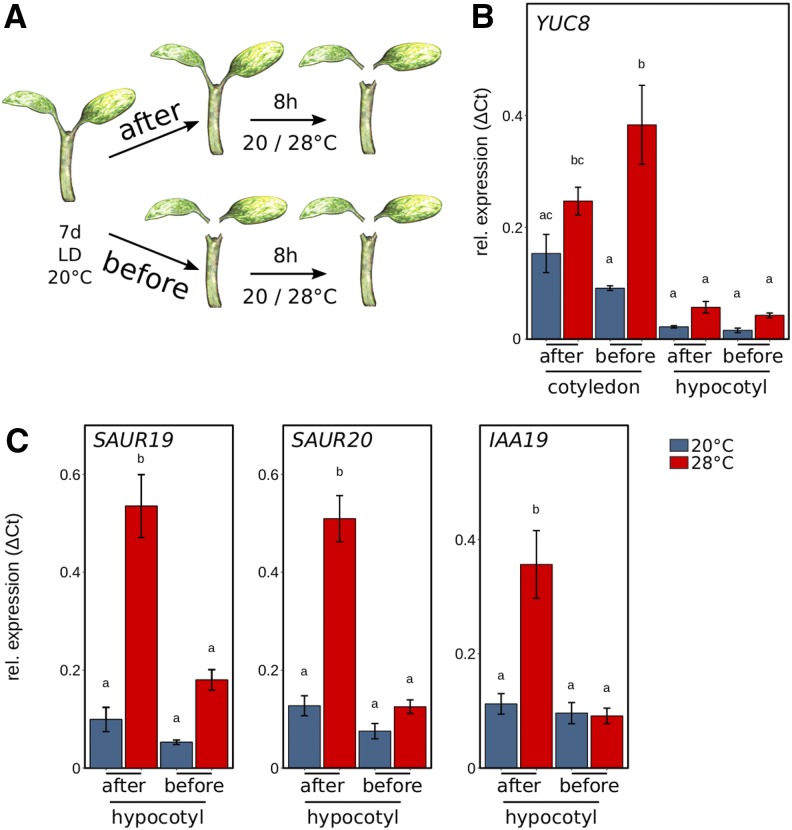
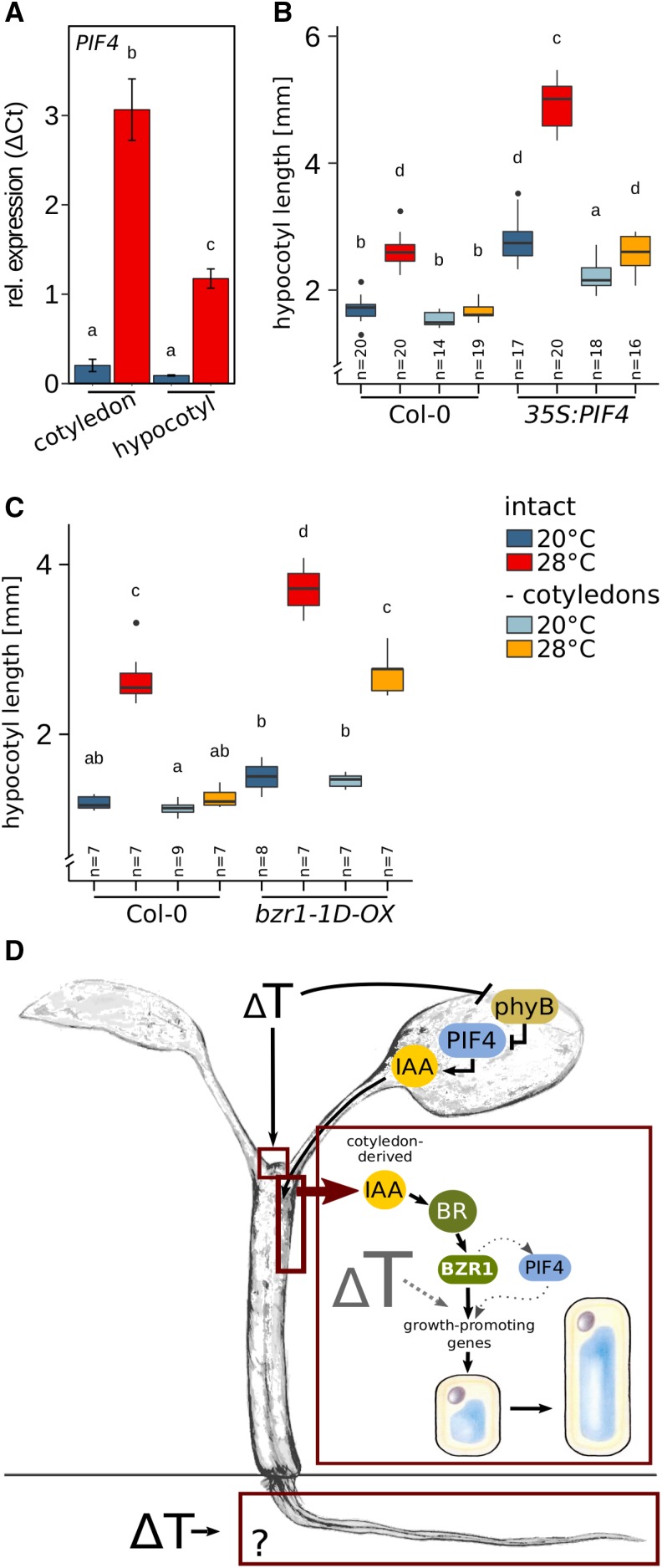
Plants have a remarkable capacity to adjust their growth and development to elevated ambient temperatures. Increased elongation growth of roots, hypocotyls, and petioles in warm temperatures are hallmarks of seedling thermomorphogenesis. In the last decade, significant progress has been made to identify the molecular signaling components regulating these growth responses. Increased ambient temperature utilizes diverse components of the light sensing and signal transduction network to trigger growth adjustments. However, it remains unknown whether temperature sensing and responses are universal processes that occur uniformly in all plant organs. Alternatively, temperature sensing may be confined to specific tissues or organs, which would require a systemic signal that mediates responses in distal parts of the plant. Here, we show that Arabidopsis (Arabidopsis thaliana) seedlings show organ-specific transcriptome responses to elevated temperatures and that thermomorphogenesis involves both autonomous and organ-interdependent temperature sensing and signaling. Seedling roots can sense and respond to temperature in a shoot-independent manner, whereas shoot temperature responses require both local and systemic processes. The induction of cell elongation in hypocotyls requires temperature sensing in cotyledons, followed by the generation of a mobile auxin signal. Subsequently, auxin travels to the hypocotyl, where it triggers local brassinosteroid-induced cell elongation in seedling stems, which depends upon a distinct, permissive temperature sensor in the hypocotyl.
© 2019 American Society of Plant Biologists. All Rights Reserved.
Figures
References
-
- Casal JJ, Balasubramanian S (2019) Thermomorphogenesis. Annu Rev Plant Biol 70: - PubMed
-
- Delker C, Sonntag L, James GV, Janitza P, Ibañez C, Ziermann H, Peterson T, Denk K, Mull S, Ziegler J, et al. (2014) The DET1-COP1-HY5 pathway constitutes a multipurpose signaling module regulating plant photomorphogenesis and thermomorphogenesis. Cell Rep 9: 1983–1989 - PubMed
-
- Delker C, van Zanten M, Quint M (2017) Thermosensing enlightened. Trends Plant Sci 22: 185–187 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
