

Estradiol-Mediated Axogenesis of Hypothalamic Neurons Requires ERK1/2 and Ryanodine Receptors-Dependent Intracellular Ca2+ Rise in Male Rats
- PMID: 31001087
- PMCID: PMC6454002
- DOI: 10.3389/fncel.2019.00122
Estradiol-Mediated Axogenesis of Hypothalamic Neurons Requires ERK1/2 and Ryanodine Receptors-Dependent Intracellular Ca2+ Rise in Male Rats
Abstract
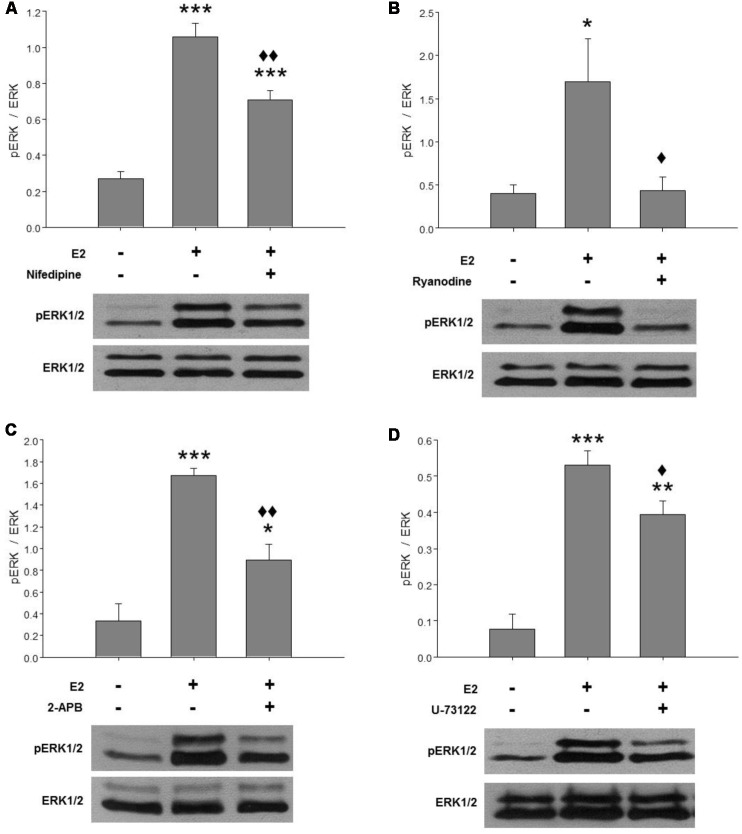
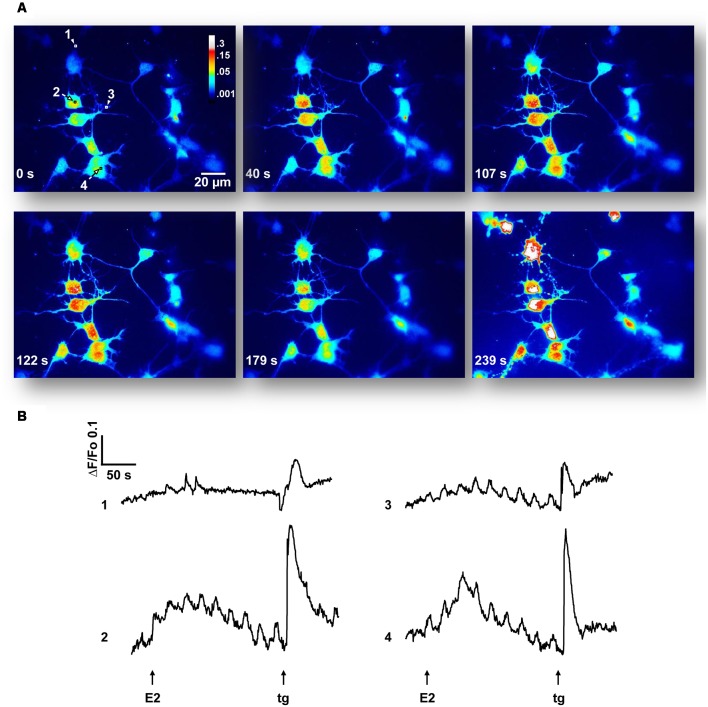
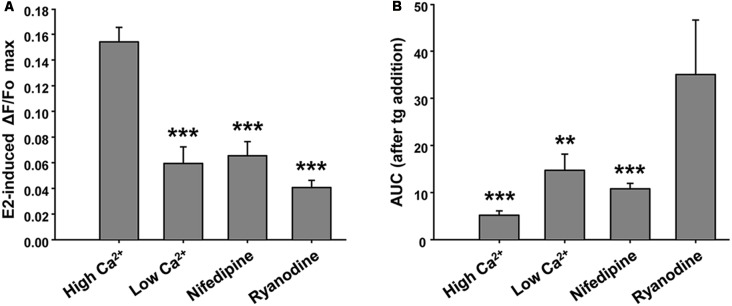
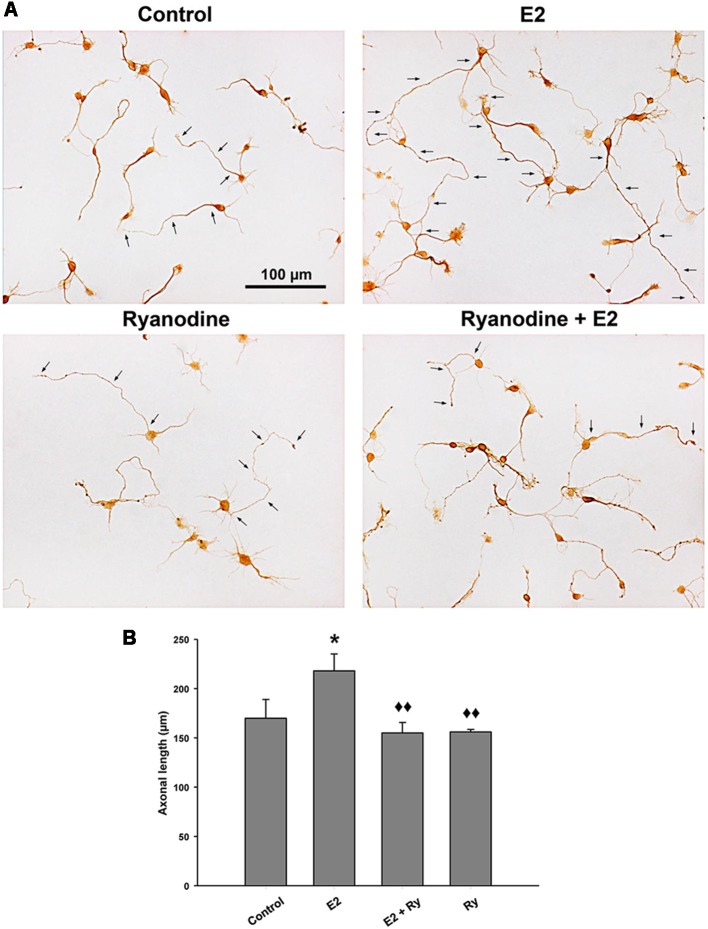
17β-estradiol (E2) induces axonal growth through extracellular signal-regulated kinase 1 and 2 (ERK1/2)-MAPK cascade in hypothalamic neurons of male rat embryos in vitro, but the mechanism that initiates these events is poorly understood. This study reports the intracellular Ca2+ increase that participates in the activation of ERK1/2 and axogenesis induced by E2. Hypothalamic neuron cultures were established from 16-day-old male rat embryos and fed with astroglia-conditioned media for 48 h. E2-induced ERK phosphorylation was completely abolished by a ryanodine receptor (RyR) inhibitor (ryanodine) and partially attenuated by an L-type voltage-gated Ca2+ channel (L-VGCC) blocker (nifedipine), an inositol-1,4,5-trisphosphate receptor (IP3R) inhibitor (2-APB), and a phospholipase C (PLC) inhibitor (U-73122). We also conducted Ca2+ imaging recording using primary cultured neurons. The results show that E2 rapidly induces an increase in cytosolic Ca2+, which often occurs in repetitive Ca2+ oscillations. This response was not observed in the absence of extracellular Ca2+ or with inhibitory ryanodine and was markedly reduced by nifedipine. E2-induced axonal growth was completely inhibited by ryanodine. In summary, the results suggest that Ca2+ mobilization from extracellular space as well as from the endoplasmic reticulum is necessary for E2-induced ERK1/2 activation and axogenesis. Understanding the mechanisms of brain estrogenic actions might contribute to develop novel estrogen-based therapies for neurodegenerative diseases.
Keywords: Ca2+ signaling; ERK1/2; axogenesis; estradiol; hypothalamic neurons; ryanodine receptors.
Figures
Similar articles
-
Axogenic effect of estrogen in male rat hypothalamic neurons involves Ca(2+), protein kinase C, and extracellular signal-regulated kinase signaling.J Neurosci Res. 2008 Jan;86(1):145-57. doi: 10.1002/jnr.21466. J Neurosci Res. 2008. PMID: 17722067
-
17Beta-estradiol induced Ca2+ influx via L-type calcium channels activates the Src/ERK/cyclic-AMP response element binding protein signal pathway and BCL-2 expression in rat hippocampal neurons: a potential initiation mechanism for estrogen-induced neuroprotection.Neuroscience. 2005;135(1):59-72. doi: 10.1016/j.neuroscience.2004.12.027. Neuroscience. 2005. PMID: 16084662
-
17beta-estradiol rapidly mobilizes intracellular calcium from ryanodine-receptor-gated stores via a PKC-PKA-Erk-dependent pathway in the human eccrine sweat gland cell line NCL-SG3.Cell Calcium. 2008 Sep;44(3):276-88. doi: 10.1016/j.ceca.2007.12.001. Epub 2008 Jan 22. Cell Calcium. 2008. PMID: 18215419
-
Thapsigargin-induced activation of Ca(2+)-CaMKII-ERK in brainstem contributes to substance P release and induction of emesis in the least shrew.Neuropharmacology. 2016 Apr;103:195-210. doi: 10.1016/j.neuropharm.2015.11.023. Epub 2015 Nov 26. Neuropharmacology. 2016. PMID: 26631534
-
Potential physiological and pathological roles for axonal ryanodine receptors.Neural Regen Res. 2023 Apr;18(4):756-759. doi: 10.4103/1673-5374.354512. Neural Regen Res. 2023. PMID: 36204832 Free PMC article. Review.
Cited by
-
Remote Ischemia Postconditioning Mitigates Hippocampal Neuron Impairment by Modulating Cav1.2-CaMKIIα-Aromatase Signaling After Global Cerebral Ischemia in Ovariectomized Rats.Mol Neurobiol. 2024 Sep;61(9):6511-6527. doi: 10.1007/s12035-024-03930-1. Epub 2024 Feb 6. Mol Neurobiol. 2024. PMID: 38321351 Free PMC article.
-
PTEN nuclear translocation enhances neuronal injury after hypoxia-ischemia via modulation of the nuclear factor-κB signaling pathway.Aging (Albany NY). 2021 Jun 10;13(12):16165-16177. doi: 10.18632/aging.203141. Epub 2021 Jun 10. Aging (Albany NY). 2021. PMID: 34114972 Free PMC article.
-
Estradiol-dependent axogenesis and Ngn3 expression are determined by XY sex chromosome complement in hypothalamic neurons.Sci Rep. 2020 May 19;10(1):8223. doi: 10.1038/s41598-020-65183-x. Sci Rep. 2020. PMID: 32427857 Free PMC article.
-
Kif21B mediates the effect of estradiol on the morphological plasticity of mouse hippocampal neurons.Front Mol Neurosci. 2023 Apr 3;16:1143024. doi: 10.3389/fnmol.2023.1143024. eCollection 2023. Front Mol Neurosci. 2023. PMID: 37078090 Free PMC article.
-
On the role of sex steroids in biological functions by classical and non-classical pathways. An update.Front Neuroendocrinol. 2021 Jul;62:100926. doi: 10.1016/j.yfrne.2021.100926. Epub 2021 Jun 3. Front Neuroendocrinol. 2021. PMID: 34089761 Free PMC article. Review.
References
-
- Adasme T., Haeger P., Paula-Lima A. C., Espinoza I., Casas-Alarcon M. M., Carrasco M. A., et al. . (2011). Involvement of ryanodine receptors in neurotrophin-induced hippocampal synaptic plasticity and spatial memory formation. Proc. Natl. Acad. Sci. U S A 108, 3029–3034. 10.1073/pnas.1013580108 - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous