

Physical and geometric determinants of transport in fetoplacental microvascular networks
- PMID: 31001587
- PMCID: PMC6469945
- DOI: 10.1126/sciadv.aav6326
Physical and geometric determinants of transport in fetoplacental microvascular networks
Abstract
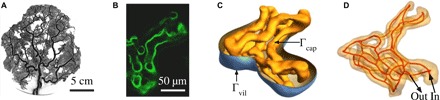
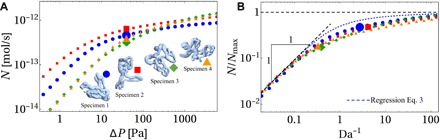
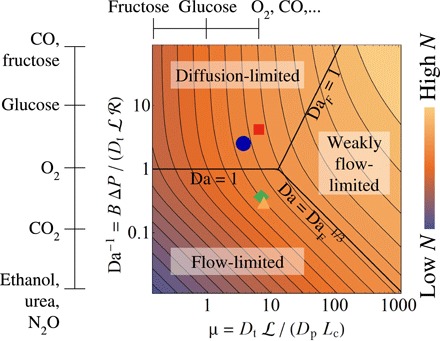
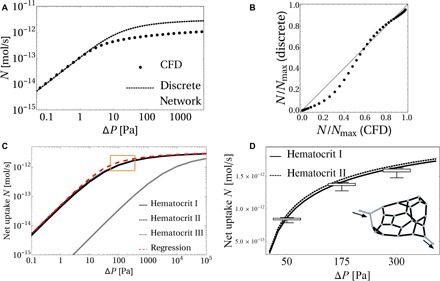
Across mammalian species, solute exchange takes place in complex microvascular networks. In the human placenta, the primary exchange units are terminal villi that contain disordered networks of fetal capillaries and are surrounded externally by maternal blood. We show how the irregular internal structure of a terminal villus determines its exchange capacity for diverse solutes. Distilling geometric features into three parameters, obtained from image analysis and computational fluid dynamics, we capture archetypal features of the structure-function relationship of terminal villi using a simple algebraic approximation, revealing transitions between flow- and diffusion-limited transport at vessel and network levels. Our theory accommodates countercurrent effects, incorporates nonlinear blood rheology, and offers an efficient method for testing network robustness. Our results show how physical estimates of solute transport, based on carefully defined geometrical statistics, provide a viable method for linking placental structure and function and offer a framework for assessing transport in other microvascular systems.
Figures

Similar articles
-
Quantifying the impact of tissue metabolism on solute transport in feto-placental microvascular networks.Interface Focus. 2019 Oct 6;9(5):20190021. doi: 10.1098/rsfs.2019.0021. Epub 2019 Aug 16. Interface Focus. 2019. PMID: 31485311 Free PMC article.
-
Modeling oxygen transport in human placental terminal villi.J Theor Biol. 2011 Dec 21;291:33-41. doi: 10.1016/j.jtbi.2011.09.008. Epub 2011 Sep 22. J Theor Biol. 2011. PMID: 21959313
-
Modelling the effect of intervillous flow on solute transfer based on 3D imaging of the human placental microstructure.Placenta. 2017 Dec;60:21-27. doi: 10.1016/j.placenta.2017.10.003. Epub 2017 Oct 12. Placenta. 2017. PMID: 29208235
-
Knowledge needed about the exchange physiology of the placenta.Placenta. 2018 Apr;64 Suppl 1:S9-S15. doi: 10.1016/j.placenta.2018.01.006. Epub 2018 Jan 19. Placenta. 2018. PMID: 29370939 Review.
-
Growth and function of the normal human placenta.Thromb Res. 2004;114(5-6):397-407. doi: 10.1016/j.thromres.2004.06.038. Thromb Res. 2004. PMID: 15507270 Review.
Cited by
-
A Graph-Based Framework for Multiscale Modeling of Physiological Transport.Front Netw Physiol. 2022 Jan 12;1:802881. doi: 10.3389/fnetp.2021.802881. eCollection 2021. Front Netw Physiol. 2022. PMID: 36925576 Free PMC article. Review.
-
ALKBH5 modulation of ferroptosis in recurrent miscarriage: implications in cytotrophoblast dysfunction.PeerJ. 2024 Oct 18;12:e18227. doi: 10.7717/peerj.18227. eCollection 2024. PeerJ. 2024. PMID: 39434797 Free PMC article.
-
Transcriptional and Epigenomic Markers of the Arterial-Venous and Micro/Macro-Vascular Endothelial Heterogeneity within the Umbilical-Placental Bed.Int J Mol Sci. 2022 Oct 6;23(19):11873. doi: 10.3390/ijms231911873. Int J Mol Sci. 2022. PMID: 36233172 Free PMC article.
-
Feature tracking microfluidic analysis reveals differential roles of viscosity and friction in sickle cell blood.Lab Chip. 2022 Apr 12;22(8):1565-1575. doi: 10.1039/d1lc01133b. Lab Chip. 2022. PMID: 35315465 Free PMC article.
-
Bioengineering Approaches for Placental Research.Ann Biomed Eng. 2021 Aug;49(8):1805-1818. doi: 10.1007/s10439-020-02714-7. Epub 2021 Jan 8. Ann Biomed Eng. 2021. PMID: 33420547 Review.
References
-
- Serov A. S., Salafia C., Grebenkov D. S., Filoche M., The role of morphology in mathematical models of placental gas exchange. J. Appl. Physiol. 120, 17–28 (2016). - PubMed
Publication types
MeSH terms
Substances
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
