

Critical role of spectrin in hearing development and deafness
- PMID: 31001589
- PMCID: PMC6469942
- DOI: 10.1126/sciadv.aav7803
Critical role of spectrin in hearing development and deafness
Abstract
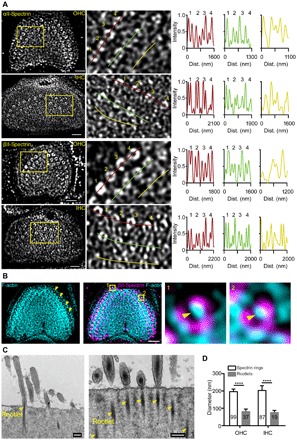
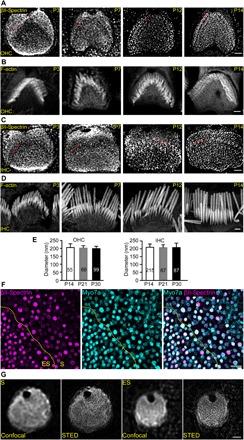
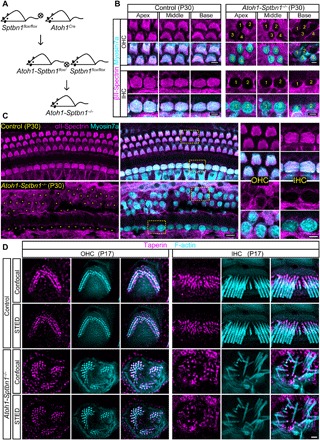
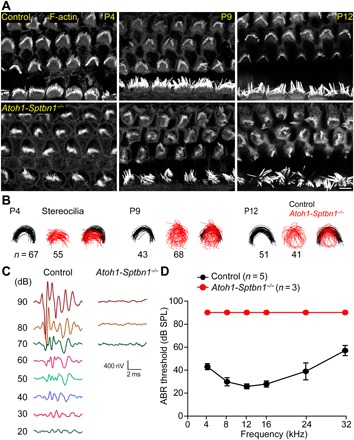
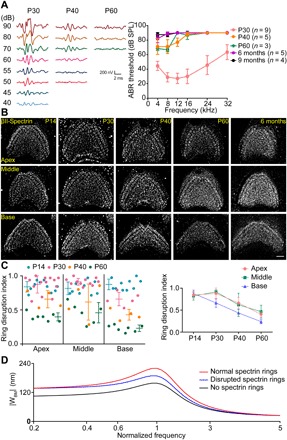
Inner ear hair cells (HCs) detect sound through the deflection of mechanosensory stereocilia. Stereocilia are inserted into the cuticular plate of HCs by parallel actin rootlets, where they convert sound-induced mechanical vibrations into electrical signals. The molecules that support these rootlets and enable them to withstand constant mechanical stresses underpin our ability to hear. However, the structures of these molecules have remained unknown. We hypothesized that αII- and βII-spectrin subunits fulfill this role, and investigated their structural organization in rodent HCs. Using super-resolution fluorescence imaging, we found that spectrin formed ring-like structures around the base of stereocilia rootlets. These spectrin rings were associated with the hearing ability of mice. Further, HC-specific, βII-spectrin knockout mice displayed profound deafness. Overall, our work has identified and characterized structures of spectrin that play a crucial role in mammalian hearing development.
Figures
References
-
- Tilney L. G., Egelman E. H., DeRosier D. J., Saunder J. C., Actin filaments, stereocilia, and hair cells of the bird cochlea. II. Packing of actin filaments in the stereocilia and in the cuticular plate and what happens to the organization when the stereocilia are bent. J. Cell Biol. 96, 822–834 (1983). - PMC - PubMed
-
- Tilney L. G., DeRosier D. J., Actin filaments, stereocilia, and hair cells of the bird cochlea: IV. How the actin filaments become organized in developing stereocilia and in the cuticular plate. Dev. Biol. 116, 119–129 (1986). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
