

Two is a Crowd: Two is a Crowd: On the Enigmatic Etiopathogenesis of Conjoined Twinning
- PMID: 31001856
- PMCID: PMC6849862
- DOI: 10.1002/ca.23387
Two is a Crowd: Two is a Crowd: On the Enigmatic Etiopathogenesis of Conjoined Twinning
Abstract
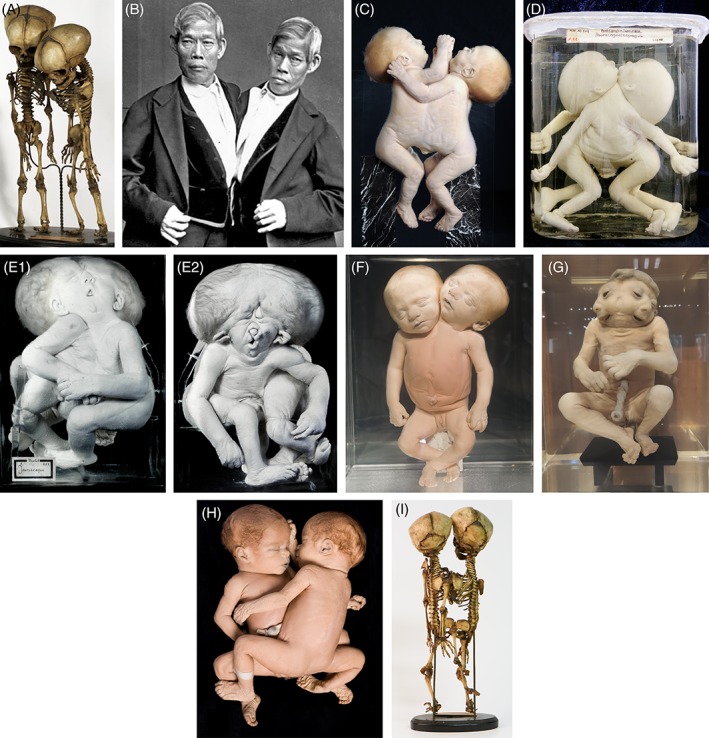
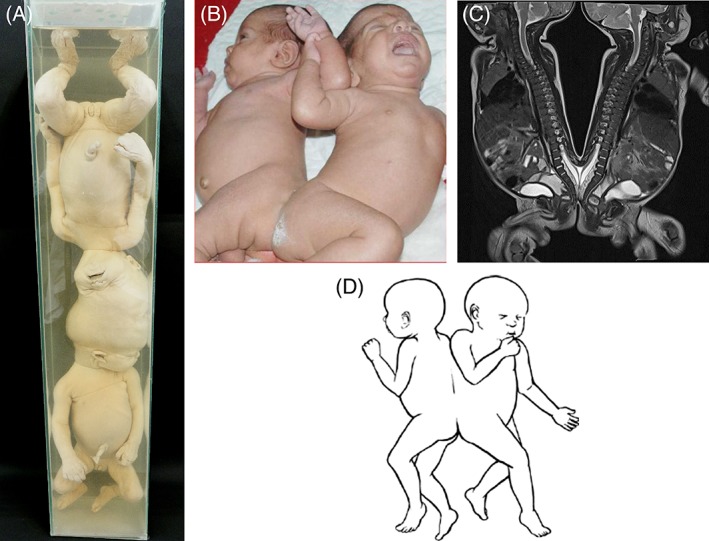
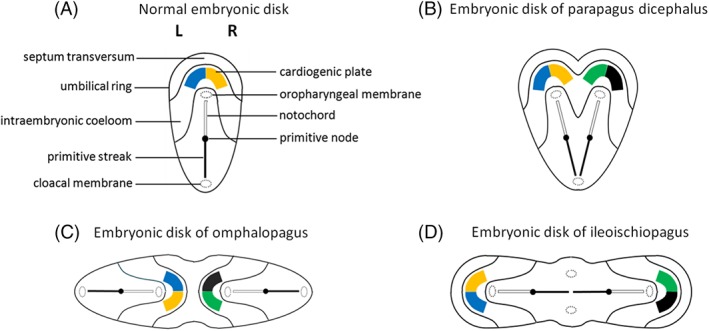
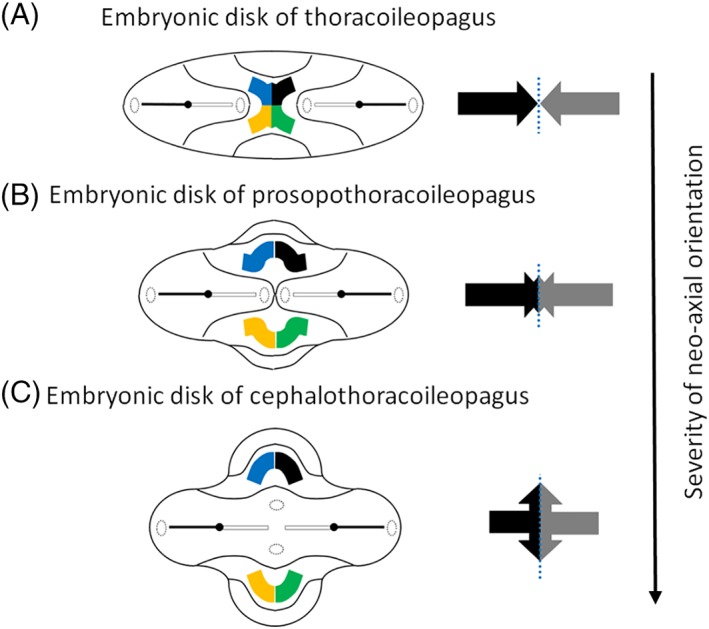
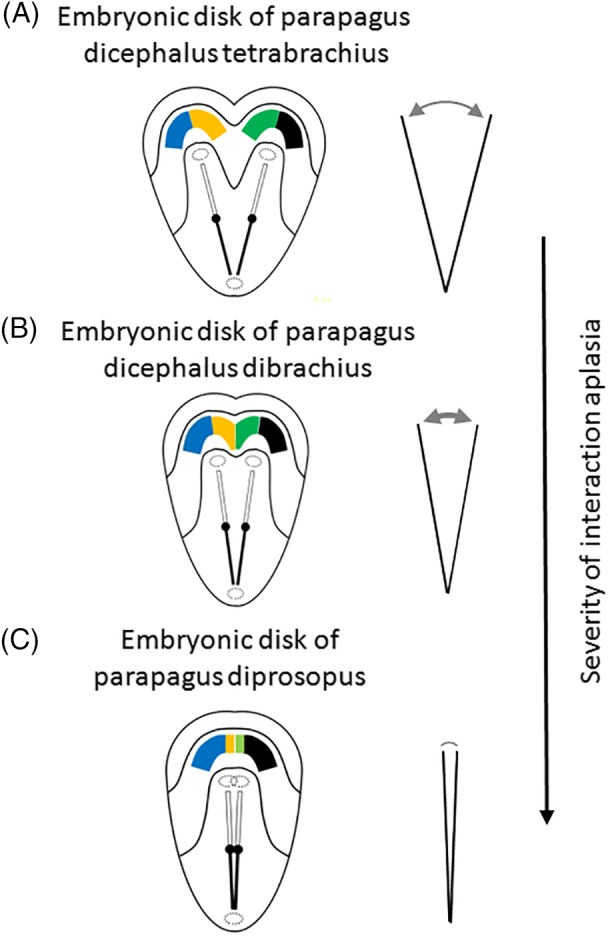
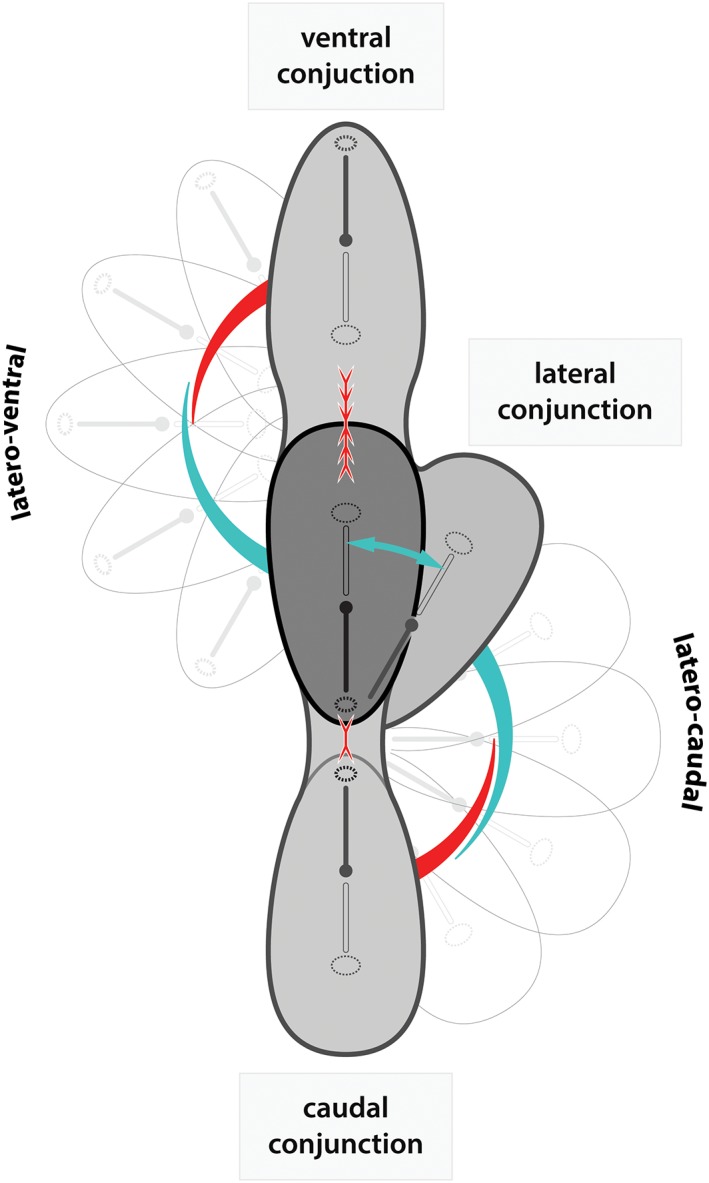
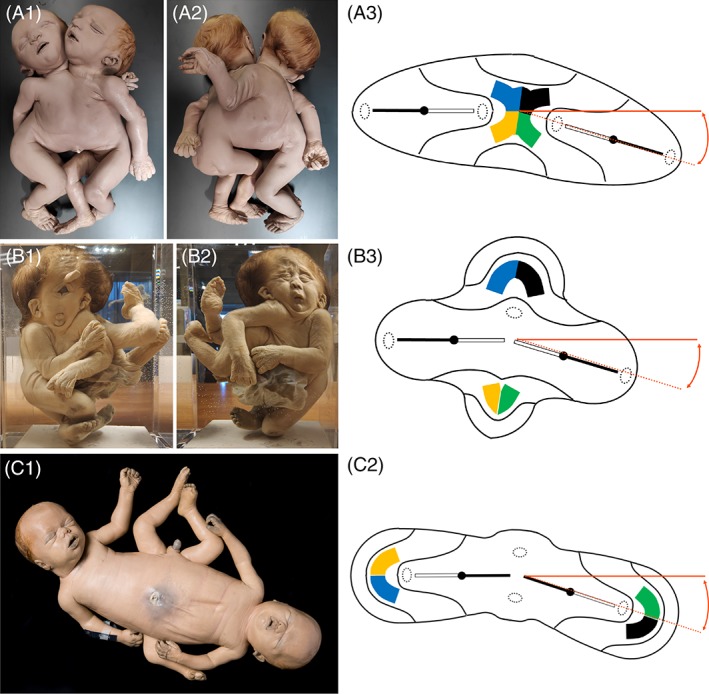
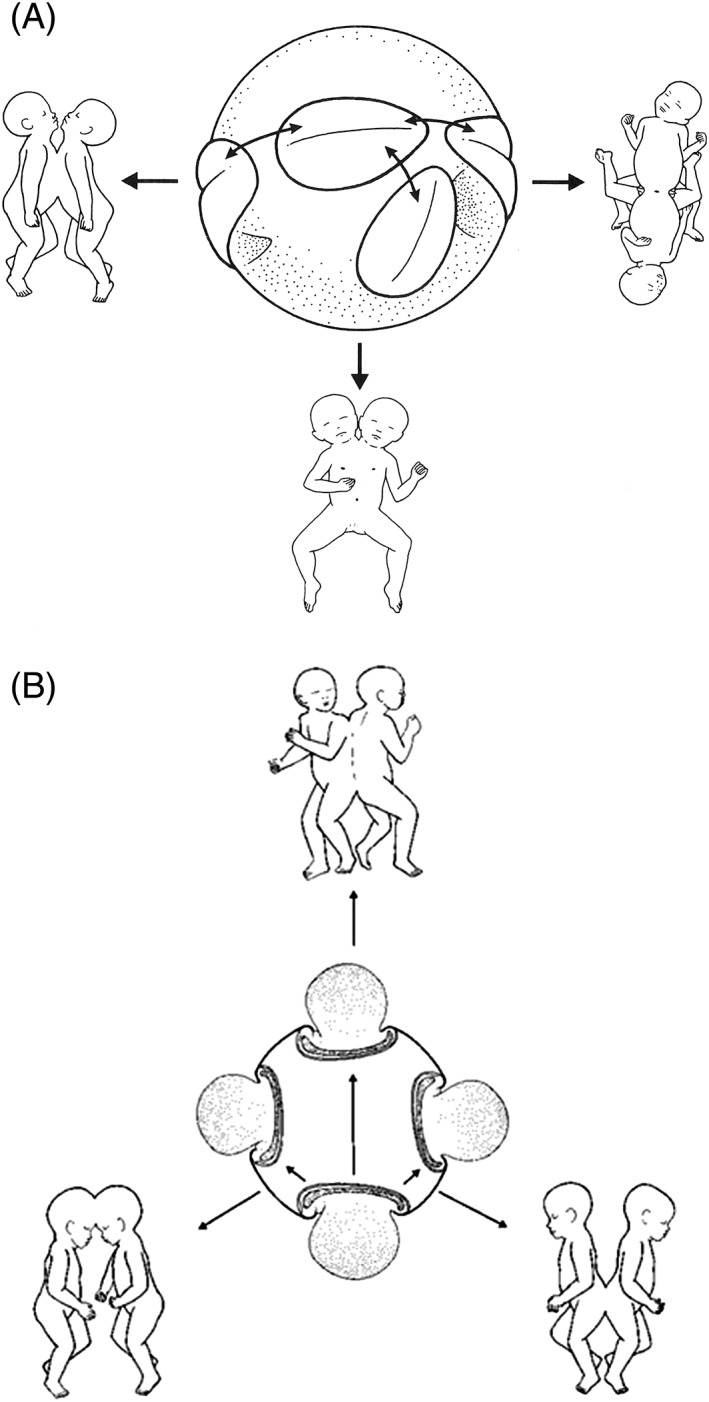
In this article, we provide a comprehensive overview of multiple facets in the puzzling genesis of symmetrical conjoined twins. The etiopathogenesis of conjoined twins remains matter for ongoing debate and is currently cited-in virtually every paper on conjoined twins-as partial fission or secondary fusion. Both theories could potentially be extrapolated from embryological adjustments exclusively seen in conjoined twins. Adoption of these, seemingly factual, theoretical proposals has (unconsciously) resulted in crystallized patterns of verbal and graphic representations concerning the enigmatic genesis of conjoined twins. Critical evaluation on their plausibility and solidity remains however largely absent. As it appears, both the fission and fusion theories cannot be applied to the full range of conjunction possibilities and thus remain matter for persistent inconclusiveness. We propose that initial duplication of axially located morphogenetic potent primordia could be the initiating factor in the genesis of ventrally, laterally, and caudally conjoined twins. The mutual position of two primordia results in neo-axial orientation and/or interaction aplasia. Both these embryological adjustments result in conjunction patterns that may seemingly appear as being caused by fission or fusion. However, as we will substantiate, neither fission nor fusion are the cause of most conjoined twinning types; rather what is interpreted as fission or fusion is actually the result of the twinning process itself. Furthermore, we will discuss the currently held views on the origin of conjoined twins and its commonly assumed etiological correlation with monozygotic twinning. Finally, considerations are presented which indicate that the dorsal conjunction group is etiologically and pathogenetically different from other symmetric conjoined twins. This leads us to propose that dorsally united twins could actually be caused by secondary fusion of two initially separate monozygotic twins. An additional reason for the ongoing etiopathogenetic debate on the genesis of conjoined twins is because different types of conjoined twins are classically placed in one overarching receptacle, which has hindered the quest for answers. Clin. Anat. 32:722-741, 2019. © 2019 Wiley Periodicals, Inc.
Keywords: cephalothoracoileopagus; conjoined twins; craniopagus; duplication; fission; fusion; ileoischiopagus; interaction aplasia; neo-axial orientation; omphalopagus; parapagus; pygopagus; rachipagus; thoracoileopagus.
© 2019 The Authors. Clinical Anatomy published by Wiley Periodicals, Inc. on behalf of American Association of Clinical Anatomists.
Figures

Similar articles
-
Conjoined twins: From conception to separation, a review.Clin Anat. 2017 Apr;30(3):385-396. doi: 10.1002/ca.22839. Clin Anat. 2017. PMID: 28195364 Review.
-
The embryology of conjoined twins.Childs Nerv Syst. 2004 Aug;20(8-9):508-25. doi: 10.1007/s00381-004-0985-4. Epub 2004 Jul 27. Childs Nerv Syst. 2004. PMID: 15278382 Review.
-
Characterizing the coalescence area of conjoined twins to elucidate congenital disorders in singletons.Clin Anat. 2021 Sep;34(6):845-858. doi: 10.1002/ca.23725. Epub 2021 Feb 20. Clin Anat. 2021. PMID: 33533057 Free PMC article.
-
Diamniotic placentation associated with omphalopagus conjoined twins: implications for a contemporary model of conjoined twinning.Am J Med Genet. 1994 Aug 15;52(2):188-95. doi: 10.1002/ajmg.1320520213. Am J Med Genet. 1994. PMID: 7802007
-
Rachipagus conjoined twins: they really do occur!Teratology. 1995 Dec;52(6):346-56. doi: 10.1002/tera.1420520605. Teratology. 1995. PMID: 8711621 Review.
Cited by
-
Phenotypically Discordant Anomalies in Conjoined Twins: Quirks of Nature Governed by Molecular Pathways?Diagnostics (Basel). 2023 Nov 10;13(22):3427. doi: 10.3390/diagnostics13223427. Diagnostics (Basel). 2023. PMID: 37998563 Free PMC article. Review.
-
Parasitic pygopagus conjoined twins' incidental findings in the obstructed labor of an intrapartum dead baby that had undergone emergency C-section: rare case report in South Papua.Childs Nerv Syst. 2024 Aug;40(8):2631-2635. doi: 10.1007/s00381-024-06440-6. Epub 2024 May 2. Childs Nerv Syst. 2024. PMID: 38695890
-
Conjoined twins and conjoined triplets: At the heart of the matter.Birth Defects Res. 2022 Jul 15;114(12):596-610. doi: 10.1002/bdr2.2066. Epub 2022 Jun 29. Birth Defects Res. 2022. PMID: 35766259 Free PMC article. Review.
-
A Detailed Description and Discussion on Conjoined Twins.Cureus. 2022 Sep 24;14(9):e29526. doi: 10.7759/cureus.29526. eCollection 2022 Sep. Cureus. 2022. PMID: 36312620 Free PMC article. Review.
-
Conjoined Twins in Guinea Pigs: A Case Report.Animals (Basel). 2022 Jul 26;12(15):1904. doi: 10.3390/ani12151904. Animals (Basel). 2022. PMID: 35892553 Free PMC article.
References
-
- Awasthi R, Iyengar R, Rege S, Jain N. 2015. Surgical management of pygopagus conjoined twins with spinal bifida. Eur Spine J 24(Suppl 4):S560–S563. - PubMed
-
- Azar Y, Eyal‐Giladi A. 1981. Interaction of epiblast and hypoblast in the formation of the primitive streak and the embryonic axis in chick, as revealed by hypoblast rotation experiments. J Embryol Exp Morphol 61:133–144. - PubMed
-
- Beckwith JB. 2002. Congenital malformations: from superstition to understanding. Virchows Arch 461:609–619. - PubMed
-
- Beddington RS. 1994. Induction of a second neural axis by the mouse node. Development 120:613–620. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials
