

Tomato fruit ripening factor NOR controls leaf senescence
- PMID: 31002305
- PMCID: PMC6506771
- DOI: 10.1093/jxb/erz098
Tomato fruit ripening factor NOR controls leaf senescence
Abstract
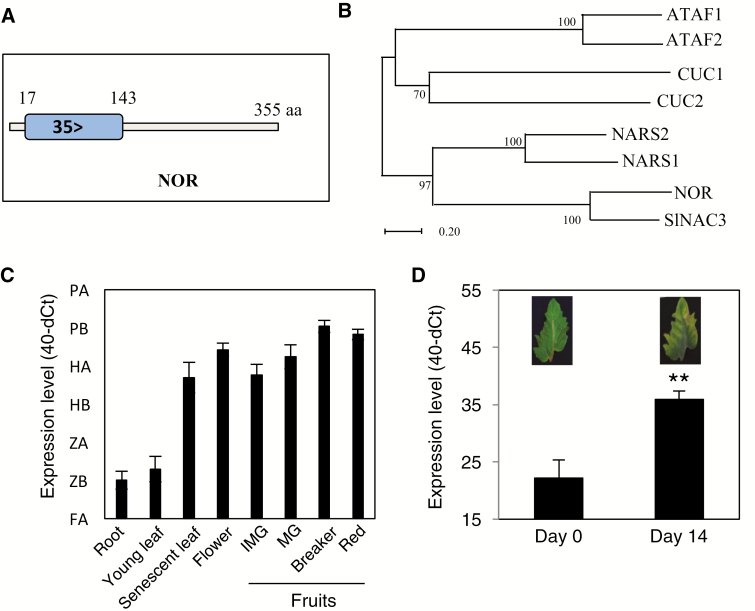
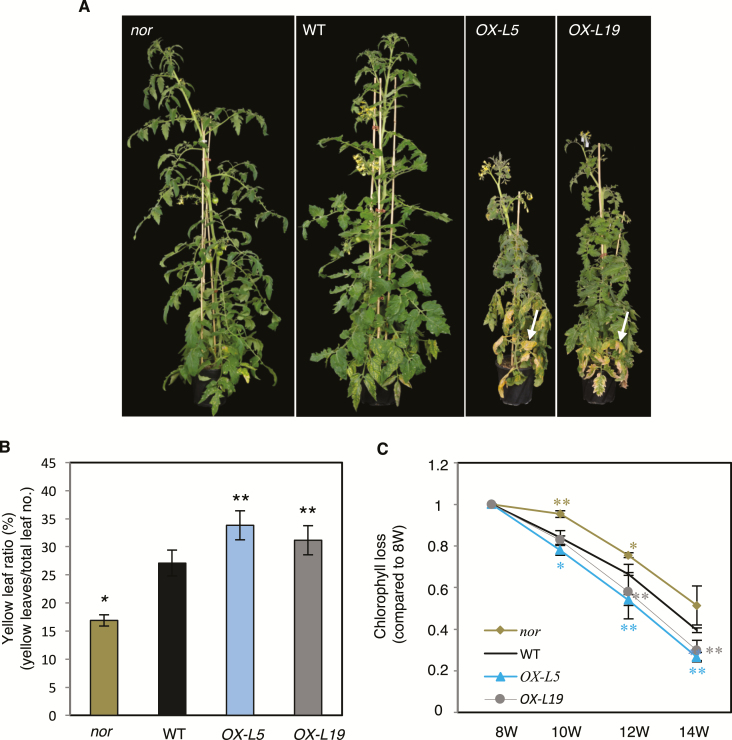
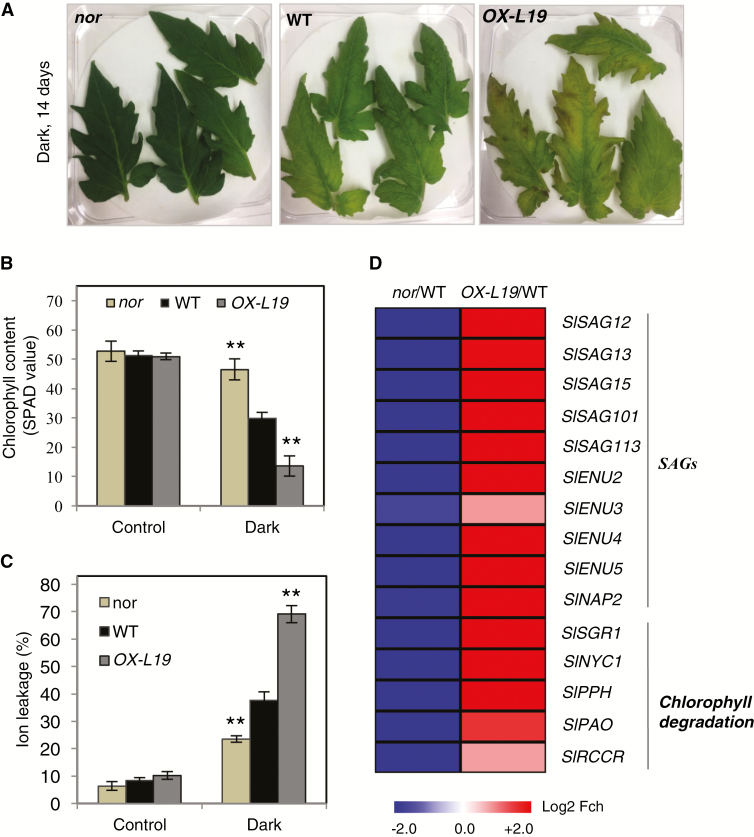
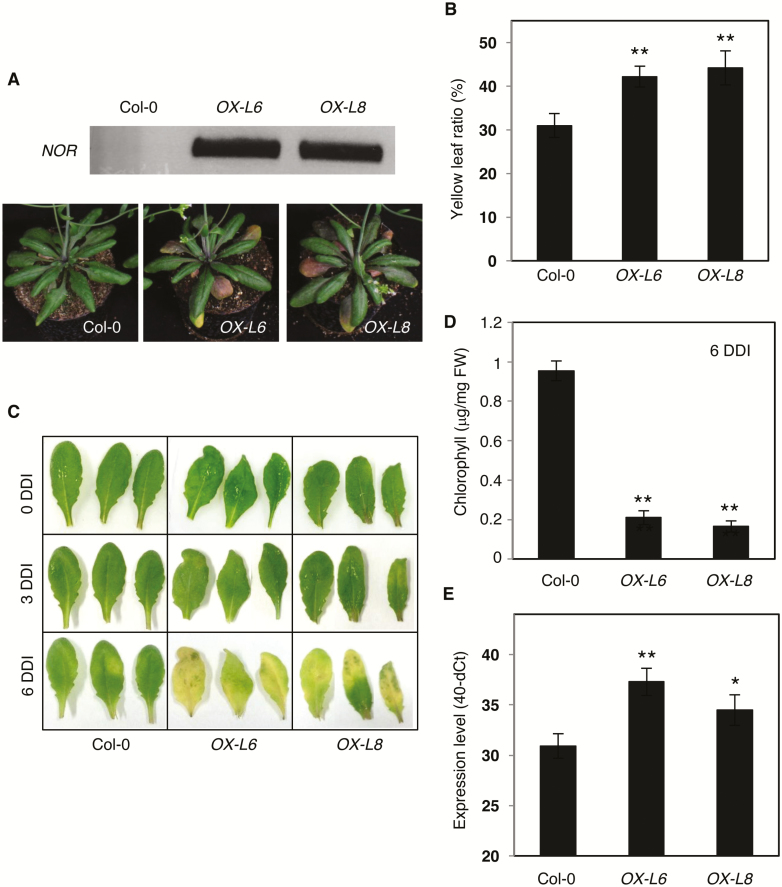
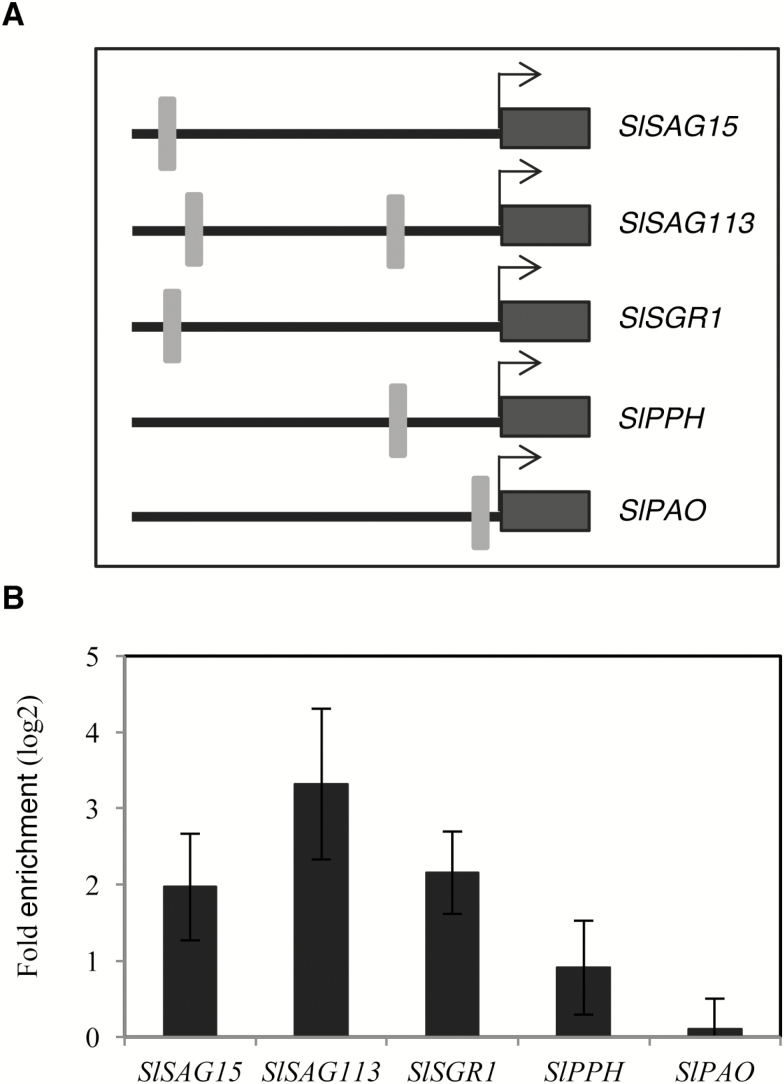
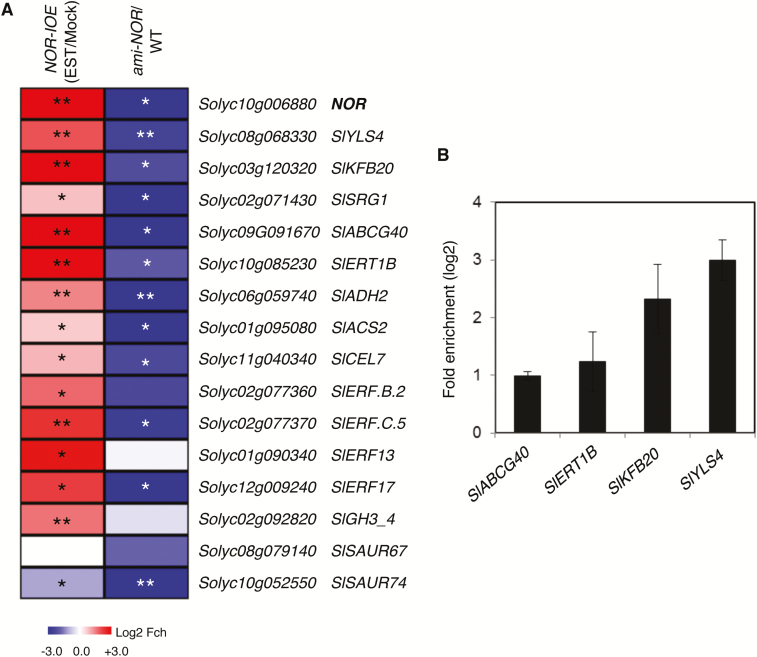
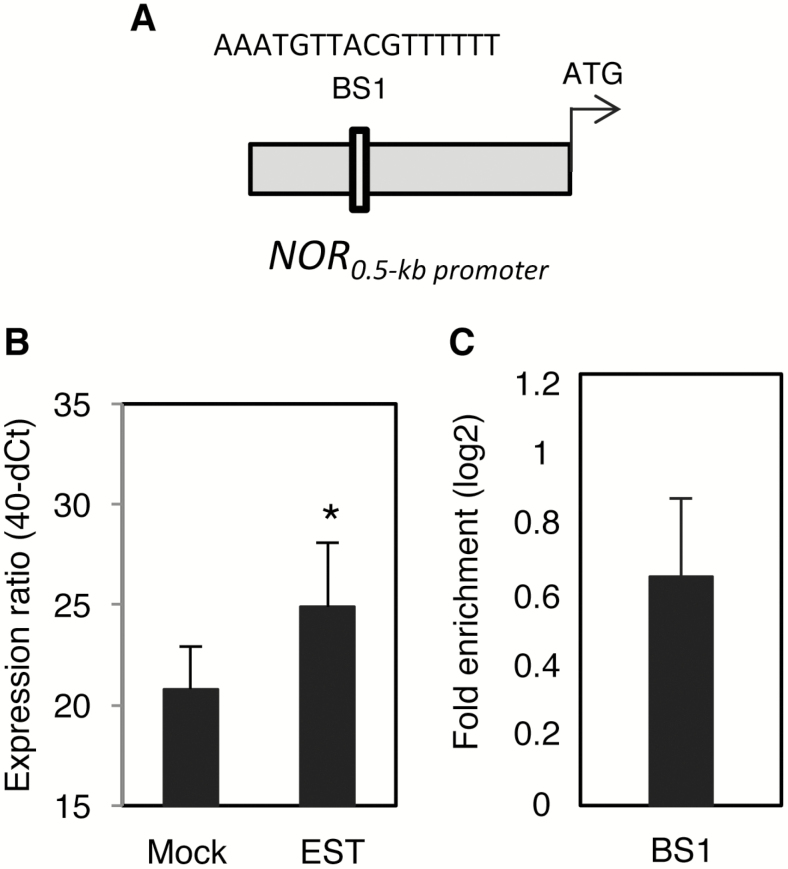
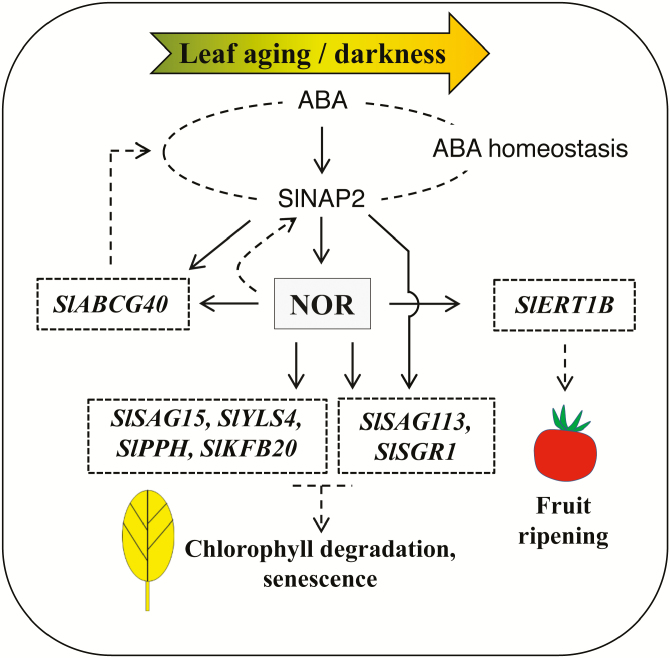
NAC transcription factors (TFs) are important regulators of expressional reprogramming during plant development, stress responses, and leaf senescence. NAC TFs also play important roles in fruit ripening. In tomato (Solanum lycopersicum), one of the best characterized NACs involved in fruit ripening is NON-RIPENING (NOR), and the non-ripening (nor) mutation has been widely used to extend fruit shelf life in elite varieties. Here, we show that NOR additionally controls leaf senescence. Expression of NOR increases with leaf age, and developmental as well as dark-induced senescence are delayed in the nor mutant, while overexpression of NOR promotes leaf senescence. Genes associated with chlorophyll degradation as well as senescence-associated genes (SAGs) show reduced and elevated expression, respectively, in nor mutants and NOR overexpressors. Overexpression of NOR also stimulates leaf senescence in Arabidopsis thaliana. In tomato, NOR supports senescence by directly and positively regulating the expression of several senescence-associated genes including, besides others, SlSAG15 and SlSAG113, SlSGR1, and SlYLS4. Finally, we find that another senescence control NAC TF, namely SlNAP2, acts upstream of NOR to regulate its expression. Our data support a model whereby NAC TFs have often been recruited by higher plants for both the control of leaf senescence and fruit ripening.
Keywords: Aging; NAC; NOR; leaf; non-ripening; senescence; tomato; transcription factor.
© The Author(s) 2019. Published by Oxford University Press on behalf of the Society for Experimental Biology.
Figures
References
-
- Balazadeh S, Riaño-Pachón DM, Mueller-Roeber B. 2008. Transcription factors regulating leaf senescence in Arabidopsis thaliana. Plant Biology 10(Suppl 1), 63–75. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
