

Structural and mechanistic themes in glycoconjugate biosynthesis at membrane interfaces
- PMID: 31003021
- PMCID: PMC6885101
- DOI: 10.1016/j.sbi.2019.03.013
Structural and mechanistic themes in glycoconjugate biosynthesis at membrane interfaces
Abstract
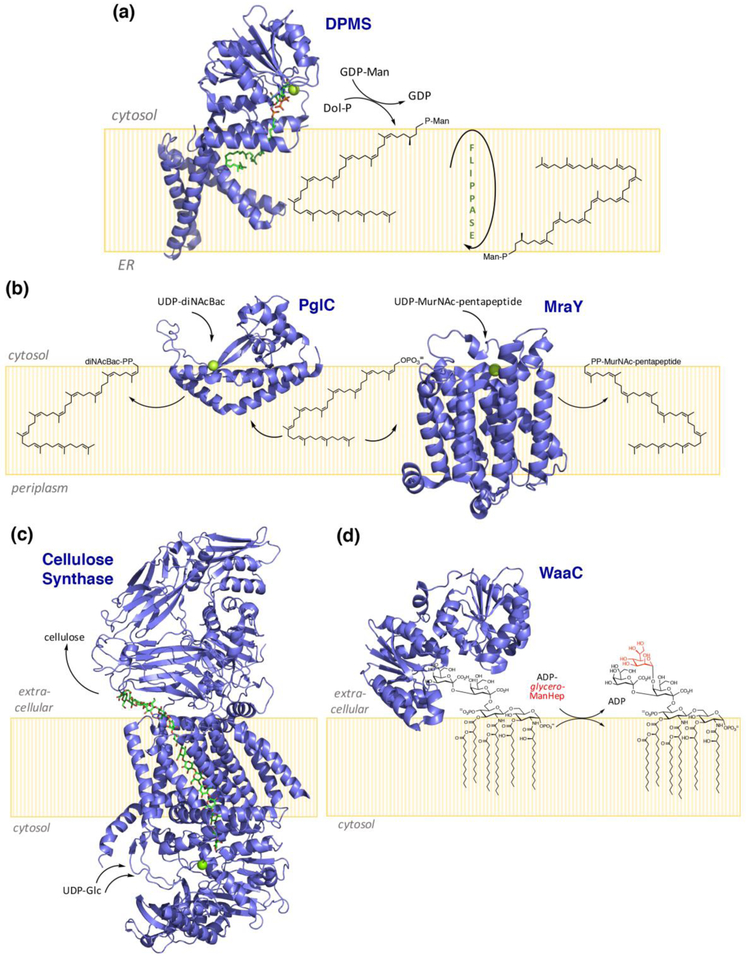
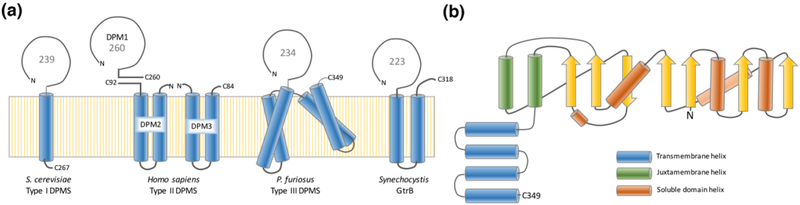
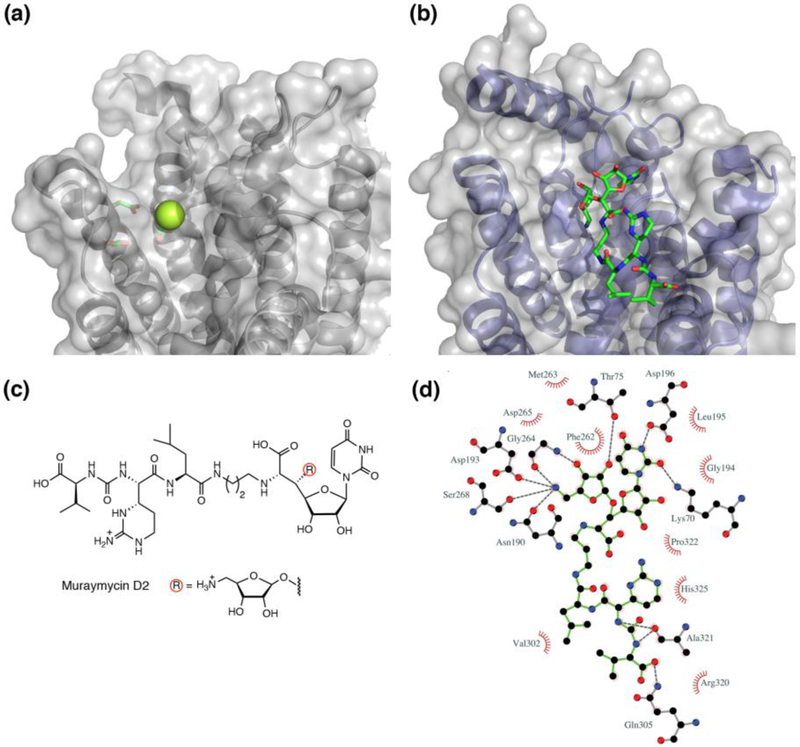
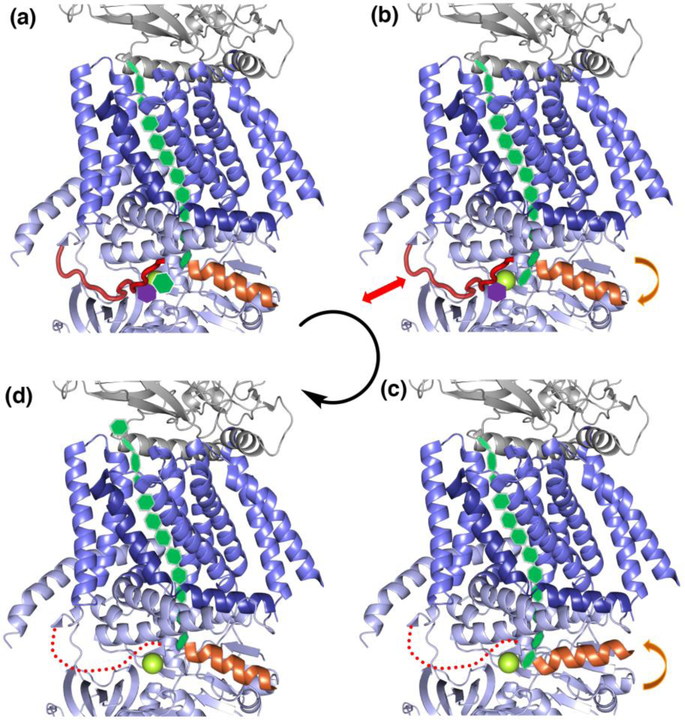
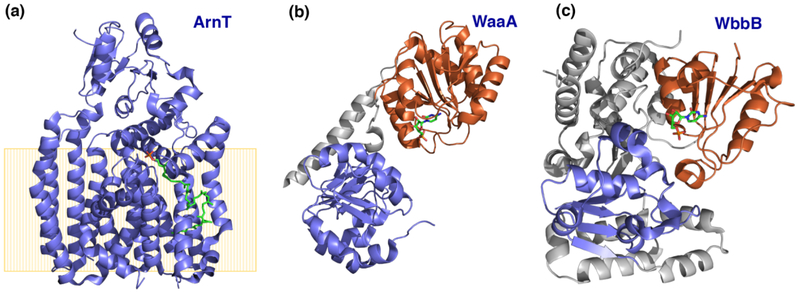
Peripheral and integral membrane proteins feature in stepwise assembly of complex glycans and glycoconjugates. Catalysis on membrane-bound substrates features challenges with substrate solubility and active-site accessibility. However, advantages in enzyme and substrate orientation and control of lateral membrane diffusion provide order to the multistep processes. Recent glycosyltransferase (GT) studies show that substrate diversity is met by the selection of folds which do not converge upon a common mechanism. Examples of polyprenol phosphate phosphoglycosyl transferases (PGTs) highlight that divergent fold families catalyze the same reaction with different mechanisms. Lipid A biosynthesis enzymes illustrate that variations on the robust Rossmann fold allow substrate diversity. Improved understanding of GT and PGT structure and function holds promise for better function prediction and improvement of therapeutic inhibitory ligands.
Copyright © 2019 Elsevier Ltd. All rights reserved.
Figures
Similar articles
-
The surprising structural and mechanistic dichotomy of membrane-associated phosphoglycosyl transferases.Biochem Soc Trans. 2021 Jun 30;49(3):1189-1203. doi: 10.1042/BST20200762. Biochem Soc Trans. 2021. PMID: 34100892 Free PMC article. Review.
-
Membrane association of monotopic phosphoglycosyl transferase underpins function.Nat Chem Biol. 2018 Jun;14(6):538-541. doi: 10.1038/s41589-018-0054-z. Epub 2018 May 16. Nat Chem Biol. 2018. PMID: 29769739 Free PMC article.
-
Bacterial phosphoglycosyl transferases: initiators of glycan biosynthesis at the membrane interface.Glycobiology. 2017 Sep 1;27(9):820-833. doi: 10.1093/glycob/cwx064. Glycobiology. 2017. PMID: 28810664 Free PMC article. Review.
-
Glycoconjugate pathway connections revealed by sequence similarity network analysis of the monotopic phosphoglycosyl transferases.Proc Natl Acad Sci U S A. 2021 Jan 26;118(4):e2018289118. doi: 10.1073/pnas.2018289118. Proc Natl Acad Sci U S A. 2021. PMID: 33472976 Free PMC article.
-
Investigation of the conserved reentrant membrane helix in the monotopic phosphoglycosyl transferase superfamily supports key molecular interactions with polyprenol phosphate substrates.Arch Biochem Biophys. 2019 Oct 30;675:108111. doi: 10.1016/j.abb.2019.108111. Epub 2019 Sep 26. Arch Biochem Biophys. 2019. PMID: 31563509 Free PMC article.
Cited by
-
The surprising structural and mechanistic dichotomy of membrane-associated phosphoglycosyl transferases.Biochem Soc Trans. 2021 Jun 30;49(3):1189-1203. doi: 10.1042/BST20200762. Biochem Soc Trans. 2021. PMID: 34100892 Free PMC article. Review.
-
Post-translational modification by the Pgf glycosylation machinery modulates Streptococcus mutans OMZ175 physiology and virulence.Mol Microbiol. 2024 Aug;122(2):133-151. doi: 10.1111/mmi.15190. Epub 2023 Nov 16. Mol Microbiol. 2024. PMID: 37972006 Free PMC article.
-
Structural basis of undecaprenyl phosphate glycosylation leading to polymyxin resistance in Gram-negative bacteria.bioRxiv [Preprint]. 2025 Jan 30:2025.01.29.634835. doi: 10.1101/2025.01.29.634835. bioRxiv. 2025. PMID: 39974898 Free PMC article. Preprint.
-
Lipoprotein N-terminal modification in Bacillus: a new paradigm for extracellular acetylation and species-dependent Toll-like receptor 2 immunomodulation.mBio. 2025 Aug 13;16(8):e0099625. doi: 10.1128/mbio.00996-25. Epub 2025 Jul 8. mBio. 2025. PMID: 40626731 Free PMC article.
-
The Bacterial Cell Wall: From Lipid II Flipping to Polymerization.Chem Rev. 2022 May 11;122(9):8884-8910. doi: 10.1021/acs.chemrev.1c00773. Epub 2022 Mar 11. Chem Rev. 2022. PMID: 35274942 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
