

M1 Muscarinic Receptors Modulate Fear-Related Inputs to the Prefrontal Cortex: Implications for Novel Treatments of Posttraumatic Stress Disorder
- PMID: 31003787
- PMCID: PMC6555658
- DOI: 10.1016/j.biopsych.2019.02.020
M1 Muscarinic Receptors Modulate Fear-Related Inputs to the Prefrontal Cortex: Implications for Novel Treatments of Posttraumatic Stress Disorder
Erratum in
-
Errata.Biol Psychiatry. 2024 Oct 15;96(8):684. doi: 10.1016/j.biopsych.2024.07.013. Epub 2024 Aug 2. Biol Psychiatry. 2024. PMID: 39093243 Free PMC article. No abstract available.
Abstract
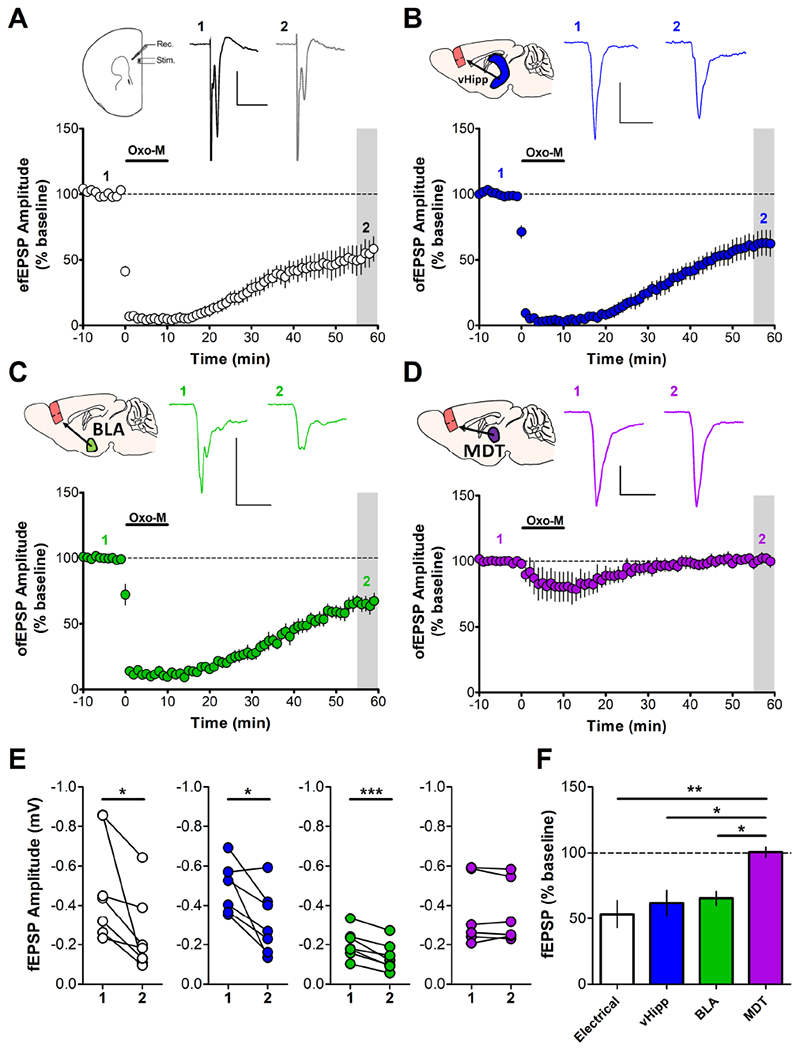
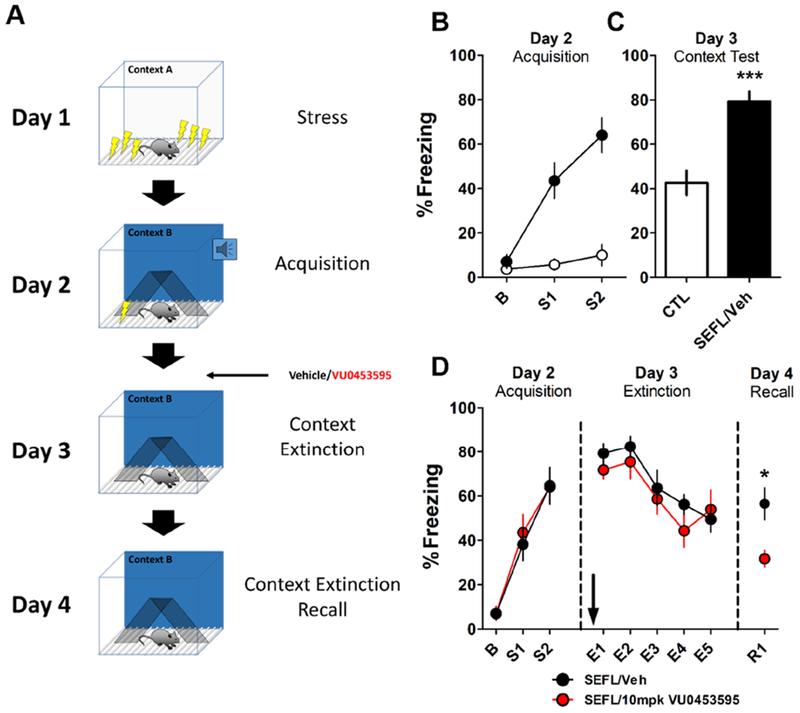
Background: The prefrontal cortex (PFC) integrates information from multiple inputs to exert top-down control allowing for appropriate responses in a given context. In psychiatric disorders such as posttraumatic stress disorder, PFC hyperactivity is associated with inappropriate fear in safe situations. We previously reported a form of muscarinic acetylcholine receptor (mAChR)-dependent long-term depression in the PFC that we hypothesize is involved in appropriate fear responding and could serve to reduce cortical hyperactivity following stress. However, it is unknown whether this long-term depression occurs at fear-related inputs.
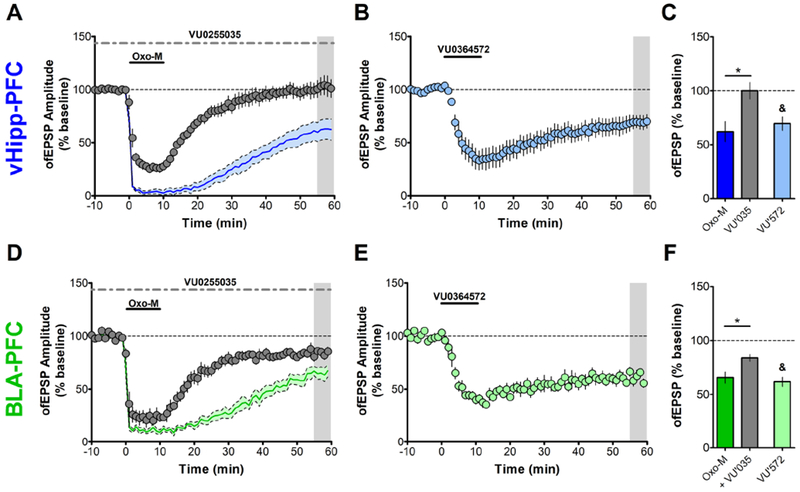
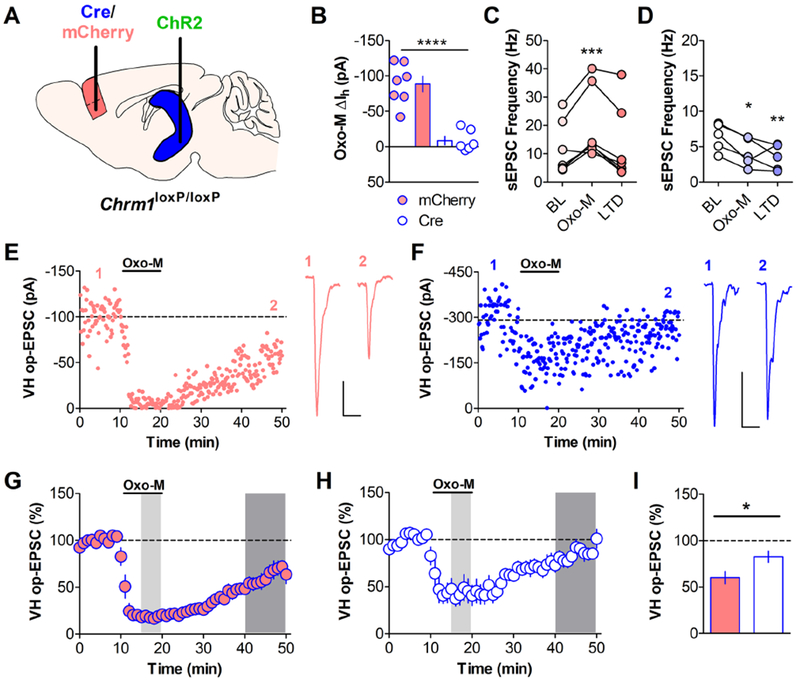
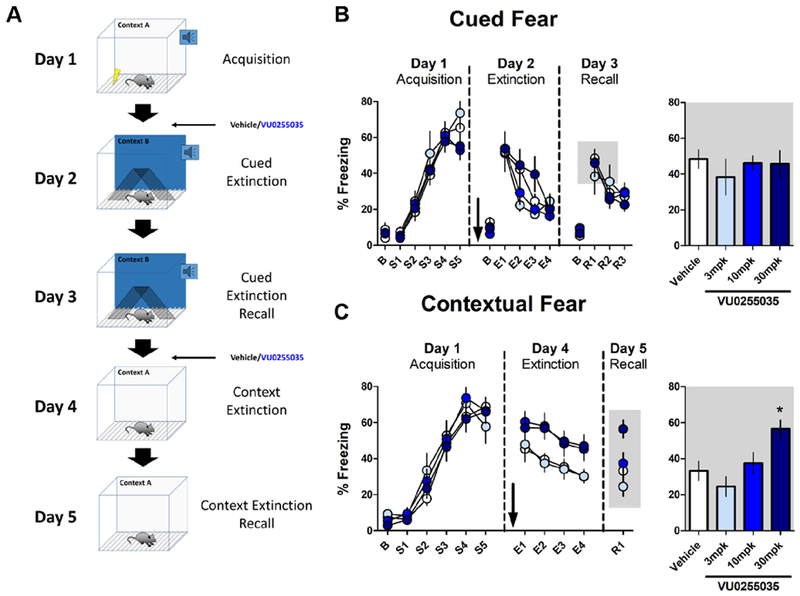
Methods: Using optogenetics with extracellular and whole-cell electrophysiology, we assessed the effect of mAChR activation on the synaptic strength of specific PFC inputs. We used selective pharmacological tools to assess the involvement of M1 mAChRs in conditioned fear extinction in control mice and in the stress-enhanced fear-learning model.
Results: M1 mAChR activation induced long-term depression at inputs from the ventral hippocampus and basolateral amygdala but not from the mediodorsal nucleus of the thalamus. We found that systemic M1 mAChR antagonism impaired contextual fear extinction. Treatment with an M1 positive allosteric modulator enhanced contextual fear extinction consolidation in stress-enhanced fear learning-conditioned mice.
Conclusions: M1 mAChRs dynamically modulate synaptic transmission at two PFC inputs whose activity is necessary for fear extinction, and M1 mAChR function is required for proper contextual fear extinction. Furthermore, an M1 positive allosteric modulator enhanced the consolidation of fear extinction in the stress-enhanced fear-learning model, suggesting that M1 positive allosteric modulators may provide a novel treatment strategy to facilitate exposure therapy in the clinic for the treatment of posttraumatic stress disorder.
Keywords: Fear extinction; M(1) muscarinic receptor; Positive allosteric modulator; Posttraumatic stress disorder; Prefrontal cortex; Synaptic plasticity.
Copyright © 2019 Society of Biological Psychiatry. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Hoover WB, Vertes RP (2007): Anatomical analysis of afferent projections to the medial prefrontal cortex in the rat. Brain Struct. Funct 212(2): 149–179. - PubMed
-
- Vertes RP (2006): Interactions among the medial prefrontal cortex, hippocampus and midline thalamus in emotional and cognitive processing in the rat. Neuroscience. 142(1): 1–20. - PubMed
-
- Goldman-Rakic PS (1995): Cellular basis of working memory. Neuron. 14(3): 477–485. - PubMed
-
- Miller EK, Cohen JD (2001): An integrative theory of prefrontal cortex function. Annu. Rev. Neurosci 24: 167–202. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
