

Three-dimensional architecture of epithelial primary cilia
- PMID: 31004057
- PMCID: PMC6511023
- DOI: 10.1073/pnas.1821064116
Three-dimensional architecture of epithelial primary cilia
Abstract
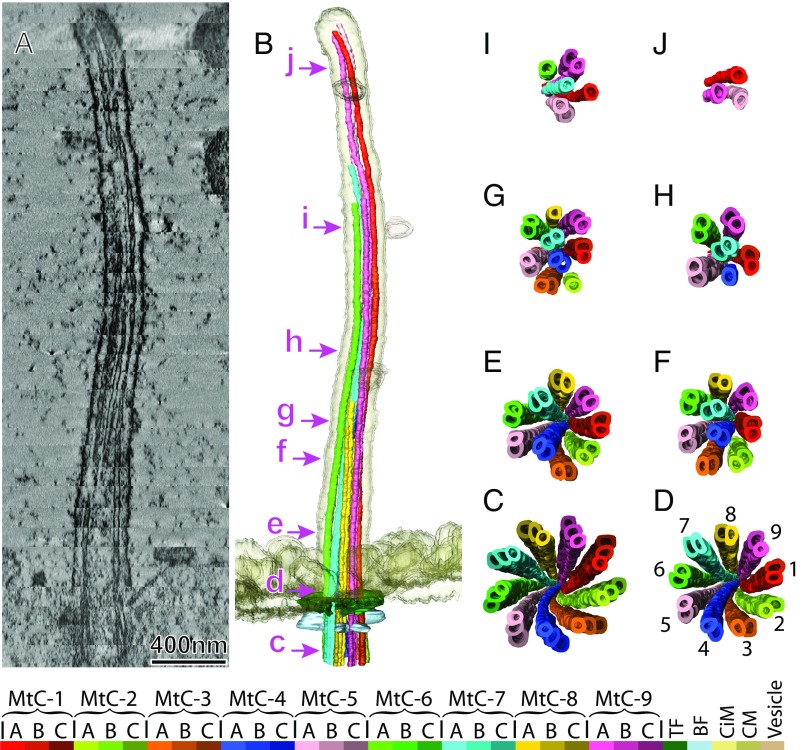
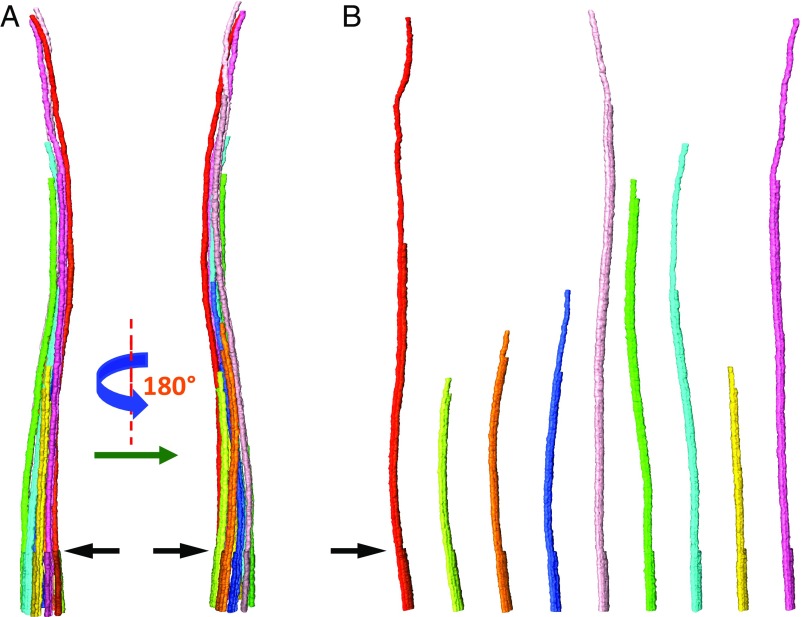
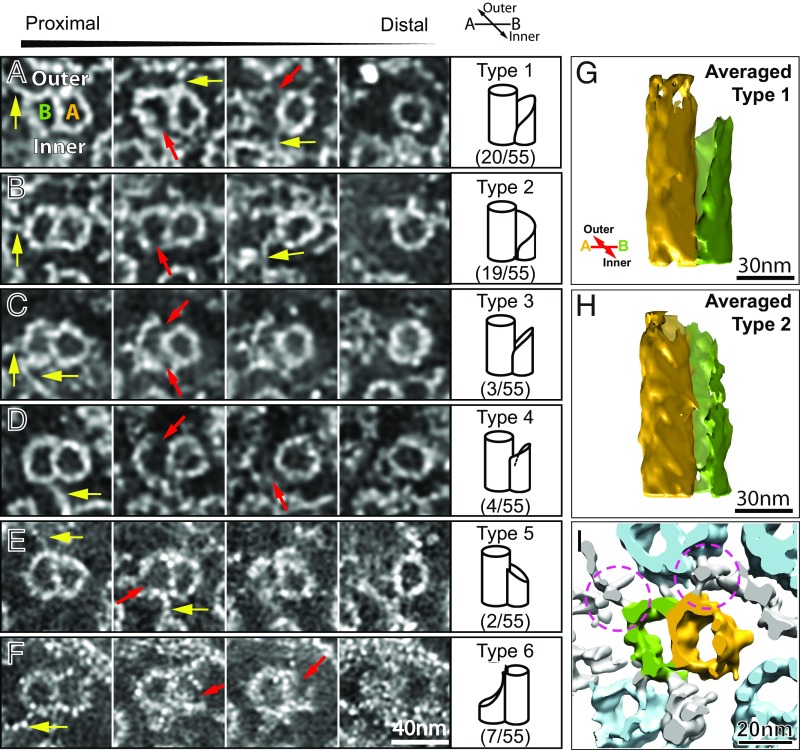
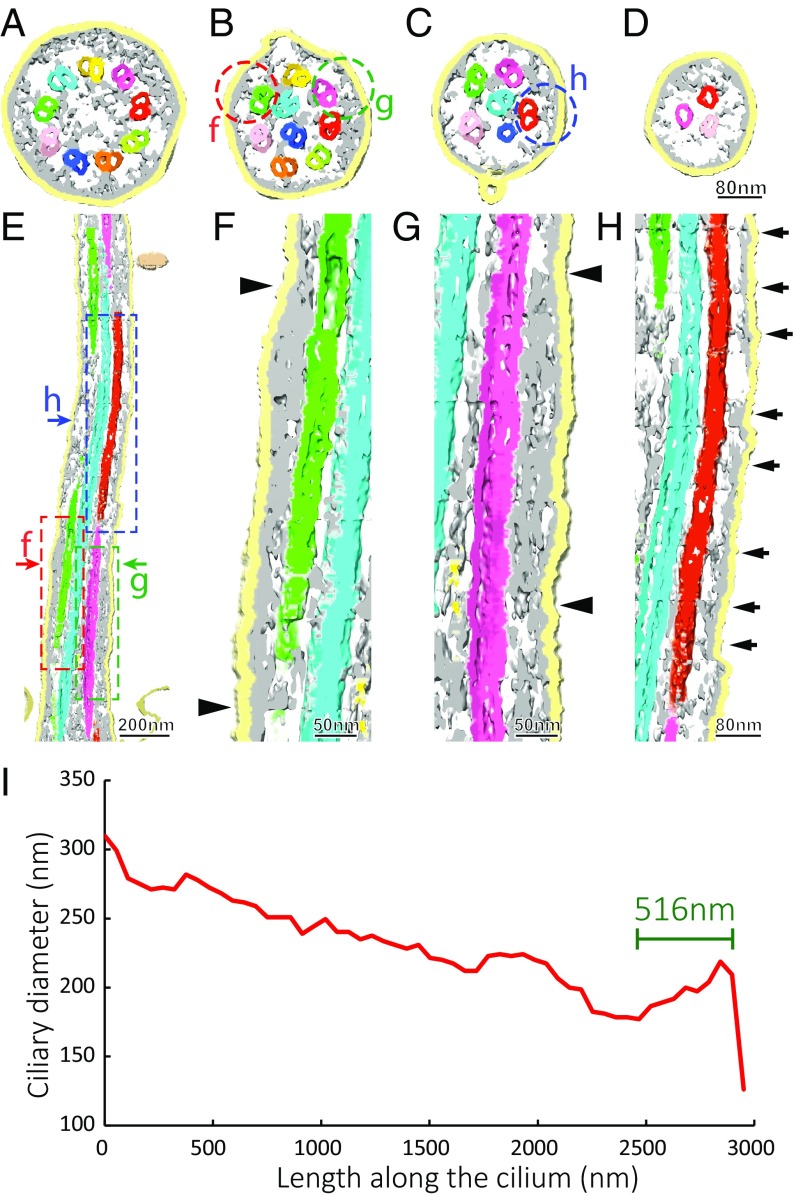
We report a complete 3D structural model of typical epithelial primary cilia based on structural maps of full-length primary cilia obtained by serial section electron tomography. Our data demonstrate the architecture of primary cilia differs extensively from the commonly acknowledged 9+0 paradigm. The axoneme structure is relatively stable but gradually evolves from base to tip with a decreasing number of microtubule complexes (MtCs) and a reducing diameter. The axonemal MtCs are cross-linked by previously unrecognized fibrous protein networks. Such an architecture explains why primary cilia can elastically withstand liquid flow for mechanosensing. The nine axonemal MtCs in a cilium are found to differ significantly in length indicating intraflagellar transport processes in primary cilia may be more complicated than that reported for motile cilia. The 3D maps of microtubule doublet-singlet transitions generally display longitudinal gaps at the inner junction between the A- and B-tubules, which indicates the inner junction protein is a major player in doublet-singlet transitions. In addition, vesicles releasing from kidney primary cilia were observed in the structural maps, supporting that ciliary vesicles budding may serve as ectosomes for cell-cell communication.
Keywords: 3D structure; electron microscopy; primary cilium.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Poole CA, Flint MH, Beaumont BW. Analysis of the morphology and function of primary cilia in connective tissues: A cellular cybernetic probe? Cell Motil. 1985;5:175–193. - PubMed
-
- Roth KE, Rieder CL, Bowser SS. Flexible-substratum technique for viewing cells from the side: Some in vivo properties of primary (9+0) cilia in cultured kidney epithelia. J Cell Sci. 1988;89:457–466. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
