

Mu opioid receptors in the medial habenula contribute to naloxone aversion
- PMID: 31005059
- PMCID: PMC6901535
- DOI: 10.1038/s41386-019-0395-7
Mu opioid receptors in the medial habenula contribute to naloxone aversion
Abstract
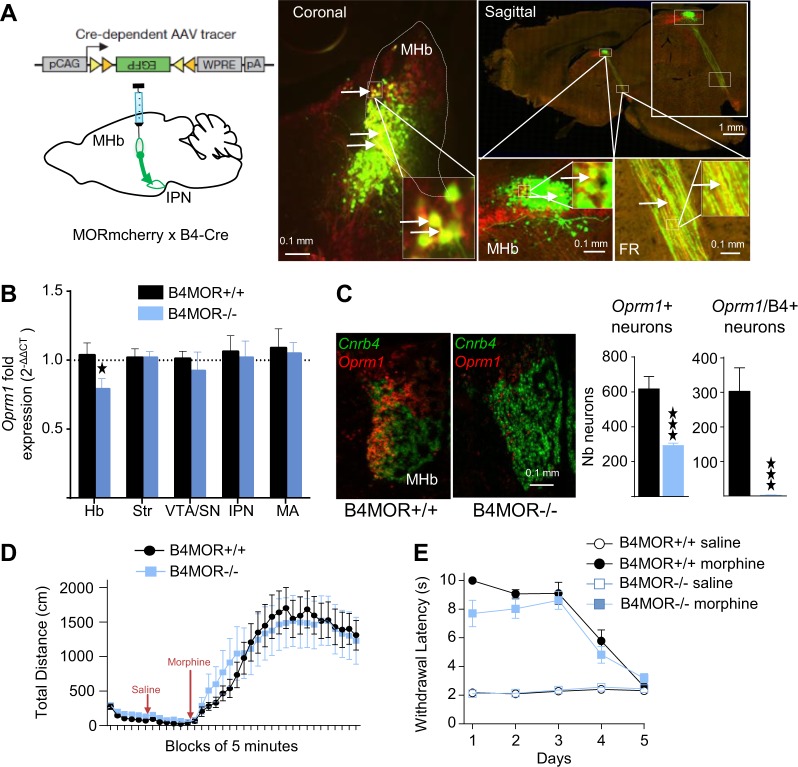
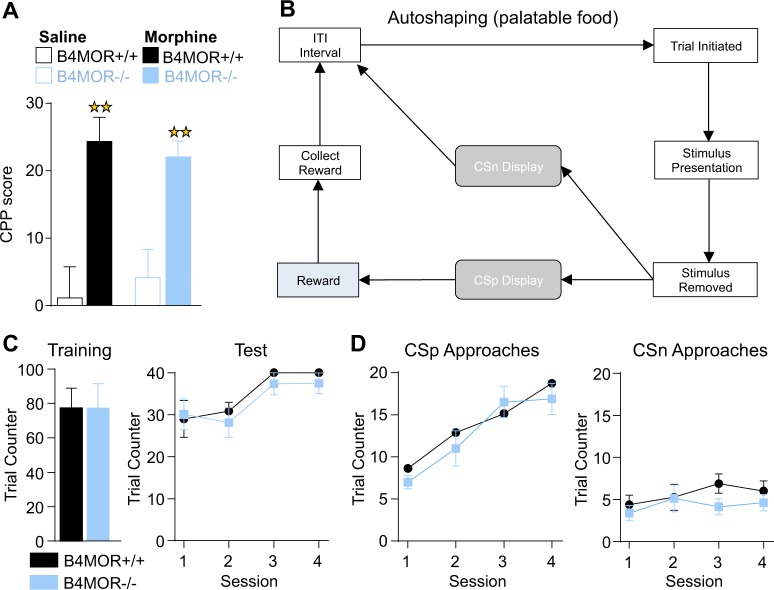
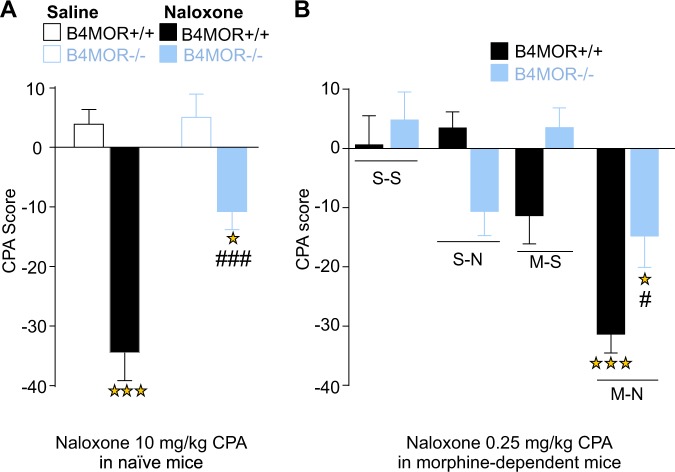
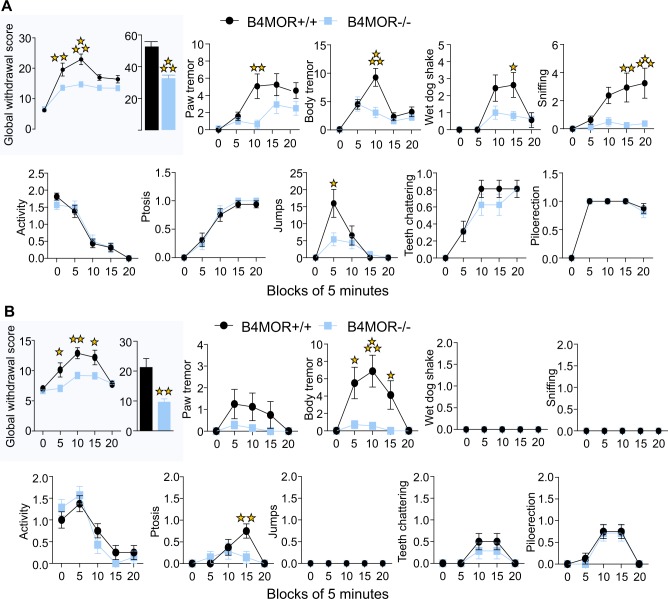
The medial habenula (MHb) is considered a brain center regulating aversive states. The mu opioid receptor (MOR) has been traditionally studied at the level of nociceptive and mesolimbic circuits, for key roles in pain relief and reward processing. MOR is also densely expressed in MHb, however, MOR function at this brain site is virtually unknown. Here we tested the hypothesis that MOR in the MHb (MHb-MOR) also regulates aversion processing. We used chnrb4-Cre driver mice to delete the Oprm1 gene in chnrb4-neurons, predominantly expressed in the MHb. Conditional mutant (B4MOR) mice showed habenula-specific reduction of MOR expression, restricted to chnrb4-neurons (50% MHb-MORs). We tested B4MOR mice in behavioral assays to evaluate effects of MOR activation by morphine, and MOR blockade by naloxone. Locomotor, analgesic, rewarding, and motivational effects of morphine were preserved in conditional mutants. In contrast, conditioned place aversion (CPA) elicited by naloxone was reduced in both naïve (high dose) and morphine-dependent (low dose) B4MOR mice. Further, physical signs of withdrawal precipitated by either MOR (naloxone) or nicotinic receptor (mecamylamine) blockade were attenuated. These data suggest that MORs expressed in MHb B4-neurons contribute to aversive effects of naloxone, including negative effect and aversive effects of opioid withdrawal. MORs are inhibitory receptors, therefore we propose that endogenous MOR signaling normally inhibits chnrb4-neurons of the MHb and moderates their known aversive activity, which is unmasked upon receptor blockade. Thus, in addition to facilitating reward at several brain sites, tonic MOR activity may also limit aversion within the MHb circuitry.
Conflict of interest statement
The authors declare no competing interest.
Figures
Similar articles
-
Expression of mu opioid receptor in dorsal diencephalic conduction system: new insights for the medial habenula.Neuroscience. 2014 Sep 26;277:595-609. doi: 10.1016/j.neuroscience.2014.07.053. Epub 2014 Jul 31. Neuroscience. 2014. PMID: 25086313 Free PMC article.
-
The mu opioid receptor and the orphan receptor GPR151 contribute to social reward in the habenula.Sci Rep. 2022 Nov 24;12(1):20234. doi: 10.1038/s41598-022-24395-z. Sci Rep. 2022. PMID: 36424418 Free PMC article.
-
Enkephalin release promotes homeostatic increases in constitutively active mu opioid receptors during morphine withdrawal.Neuroscience. 2007 Nov 9;149(3):642-9. doi: 10.1016/j.neuroscience.2007.05.011. Epub 2007 Oct 1. Neuroscience. 2007. PMID: 17905519
-
Basal opioid receptor activity, neutral antagonists, and therapeutic opportunities.Life Sci. 2005 Feb 11;76(13):1427-37. doi: 10.1016/j.lfs.2004.10.024. Epub 2004 Dec 8. Life Sci. 2005. PMID: 15680308 Review.
-
From controlled to compulsive drug-taking: The role of the habenula in addiction.Neurosci Biobehav Rev. 2019 Nov;106:102-111. doi: 10.1016/j.neubiorev.2018.06.018. Epub 2018 Jun 21. Neurosci Biobehav Rev. 2019. PMID: 29936111 Free PMC article. Review.
Cited by
-
Habenular molecular targets for depression, impulsivity, and addiction.Expert Opin Ther Targets. 2023 Jul-Dec;27(9):757-761. doi: 10.1080/14728222.2023.2257390. Epub 2023 Sep 14. Expert Opin Ther Targets. 2023. PMID: 37705488 Free PMC article. No abstract available.
-
Understanding the habenula: A major node in circuits regulating emotion and motivation.Pharmacol Res. 2023 Apr;190:106734. doi: 10.1016/j.phrs.2023.106734. Epub 2023 Mar 16. Pharmacol Res. 2023. PMID: 36933754 Free PMC article. Review.
-
Perigestational Opioid Exposure Alters Alcohol-Driven Reward Behaviors in Adolescent Rats.bioRxiv [Preprint]. 2023 Nov 16:2023.11.14.567041. doi: 10.1101/2023.11.14.567041. bioRxiv. 2023. PMID: 38014019 Free PMC article. Preprint.
-
Neuronal Excitability in the Medial Habenula and Ventral Tegmental Area Is Differentially Modulated by Nicotine Dosage and Menthol in a Sex-Specific Manner.eNeuro. 2024 Feb 12;11(2):ENEURO.0380-23.2024. doi: 10.1523/ENEURO.0380-23.2024. Print 2024 Feb. eNeuro. 2024. PMID: 38233142 Free PMC article.
-
Opioid system is necessary but not sufficient for antidepressive actions of ketamine in rodents.Proc Natl Acad Sci U S A. 2020 Feb 4;117(5):2656-2662. doi: 10.1073/pnas.1916570117. Epub 2020 Jan 15. Proc Natl Acad Sci U S A. 2020. PMID: 31941713 Free PMC article.
References
-
- Soria-Gomez E, Busquets-Garcia A, Hu F, Mehidi A, Cannich A, Roux L, et al. Habenular CB1 receptors control the expression of aversive memories. Neuron. 2015;88:306–13. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
