

Heterogeneity and emergent behaviour in the vascular endothelium
- PMID: 31005824
- PMCID: PMC6700393
- DOI: 10.1016/j.coph.2019.03.008
Heterogeneity and emergent behaviour in the vascular endothelium
Abstract
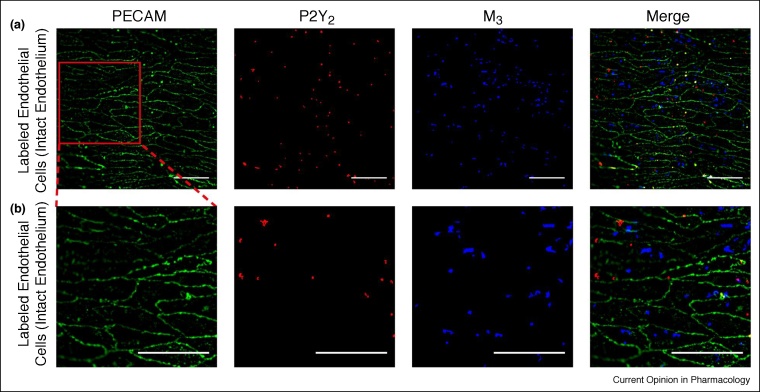
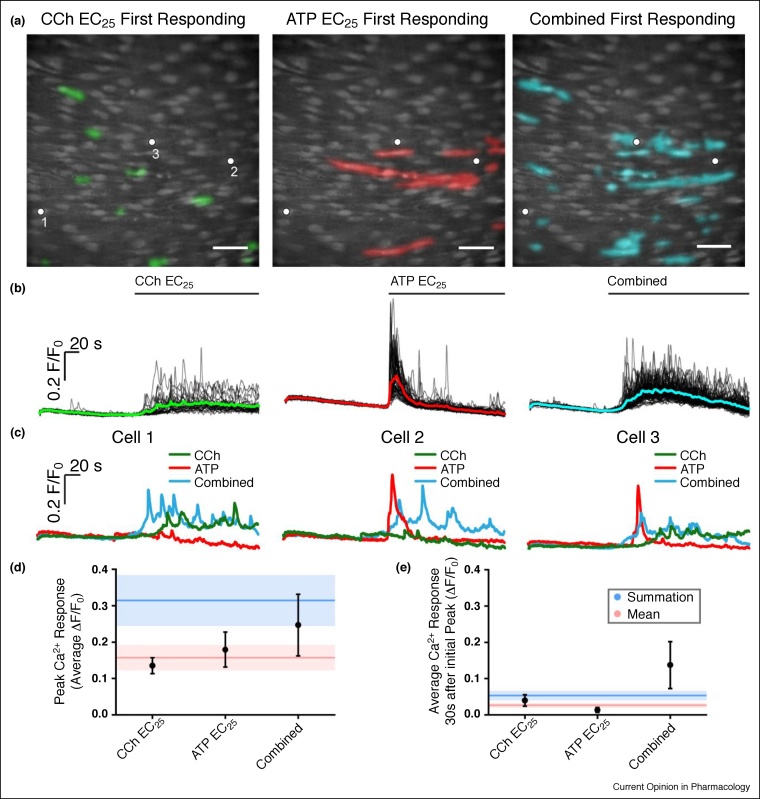
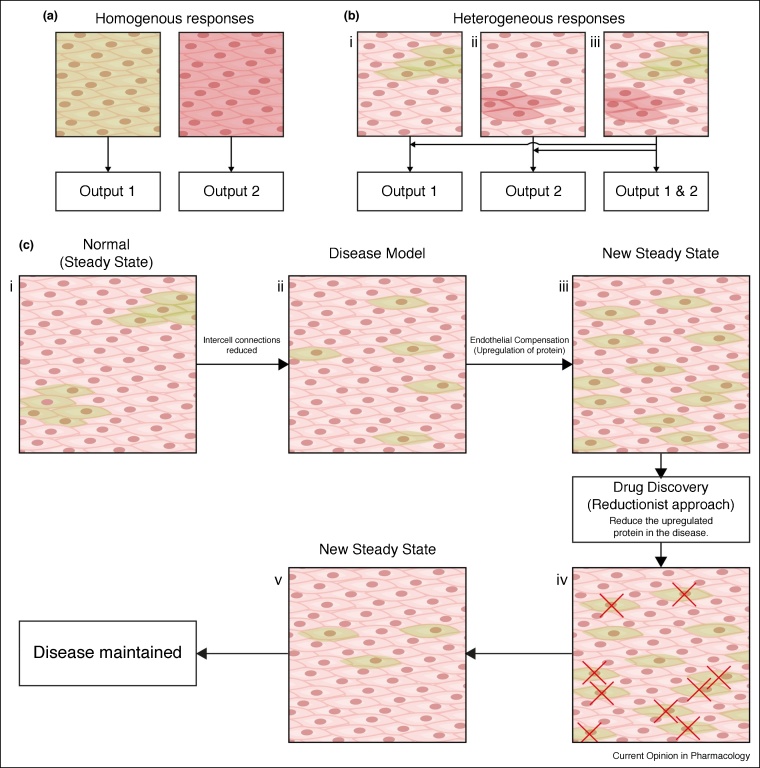
The endothelium is the single layer of cells lining all blood vessels, and it is a remarkable cardiovascular control centre. Each endothelial cell has only a small number (on average six) of interconnected neighbours. Yet this arrangement produces a large repertoire of behaviours, capable of controlling numerous cardiovascular functions in a flexible and dynamic way. The endothelium regulates the delivery of nutrients and removal of waste by regulating blood flow and vascular permeability. The endothelium regulates blood clotting, responses to infection and inflammation, the formation of new blood vessels, and remodelling of the blood vessel wall. To carry out these roles, the endothelium autonomously interprets a complex environment crammed with signals from hormones, neurotransmitters, pericytes, smooth muscle cells, various blood cells, viral or bacterial infection and proinflammatory cytokines. It is generally assumed that the endothelium responds to these instructions with coordinated responses in a homogeneous population of endothelial cells. Here, we highlight evidence that shows that neighbouring endothelial cells are highly heterogeneous and display different sensitivities to various activators. Cells with various sensitivities process different extracellular signals into distinct streams of information in parallel, like a vast switchboard. Communication occurs among cells and new 'emergent' signals are generated that are non-linear composites of the inputs. Emergent signals cannot be predicted or deduced from the properties of individual cells. Heterogeneity and emergent behaviour bestow capabilities on the endothelial collective that far exceed those of individual cells. The implications of heterogeneity and emergent behaviour for understanding vascular disease and drug discovery are discussed.
Copyright © 2019 The Authors. Published by Elsevier Ltd.. All rights reserved.
Figures
References
-
- Fraisl P., Mazzone M., Schmidt T., Carmeliet P. Regulation of angiogenesis by oxygen and metabolism. Dev Cell. 2009;16:167–179. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
