

Conformational Dynamics and Allosteric Regulation Landscapes of Germline PTEN Mutations Associated with Autism Compared to Those Associated with Cancer
- PMID: 31006514
- PMCID: PMC6506791
- DOI: 10.1016/j.ajhg.2019.03.009
Conformational Dynamics and Allosteric Regulation Landscapes of Germline PTEN Mutations Associated with Autism Compared to Those Associated with Cancer
Abstract
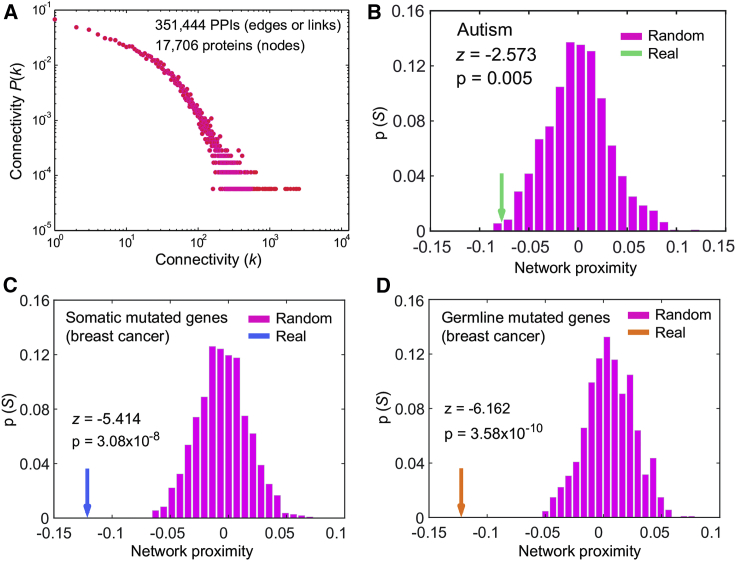
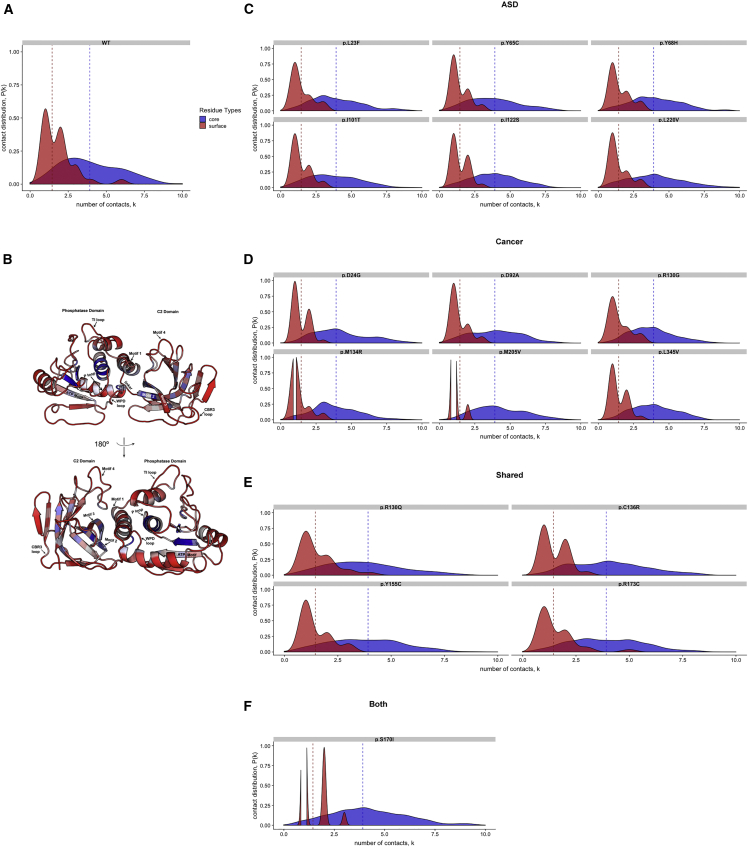
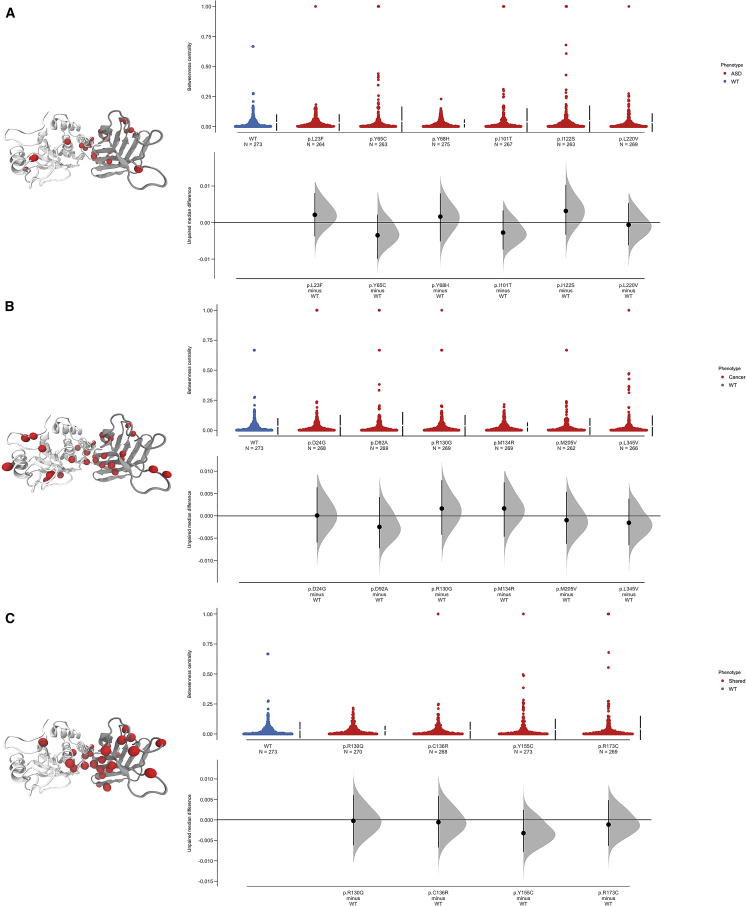
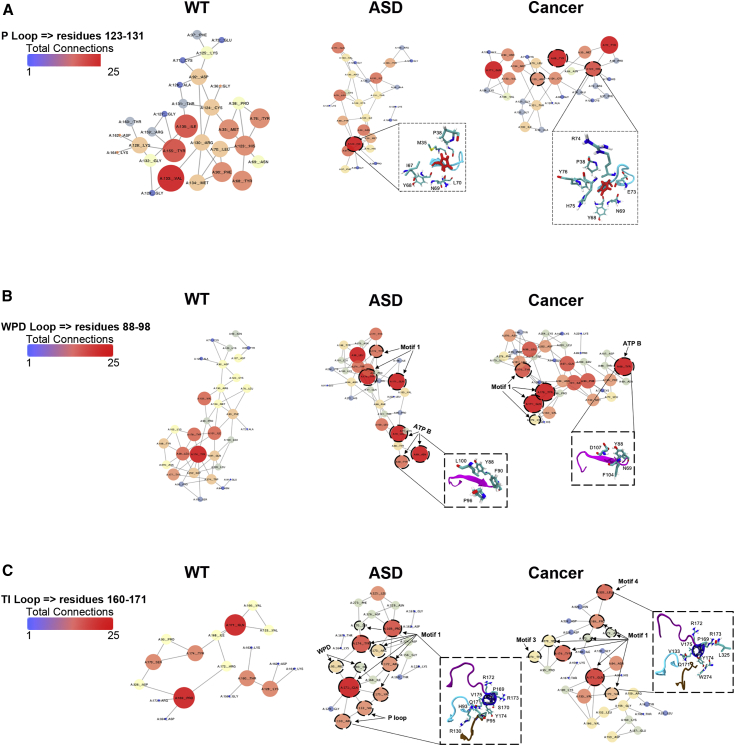
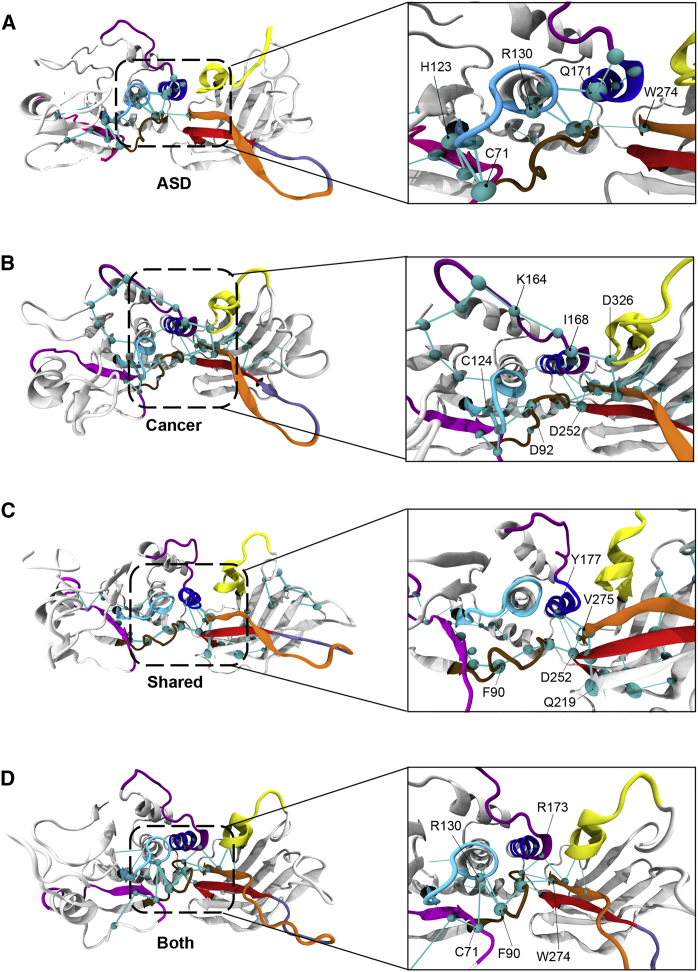
Individuals with germline PTEN tumor-suppressor variants have PTEN hamartoma tumor syndrome (PHTS). Clinically, PHTS has variable presentations; there are distinct subsets of PHTS-affected individuals, such as those diagnosed with autism spectrum disorder (ASD) or cancer. It remains unclear why mutations in one gene can lead to such seemingly disparate phenotypes. Therefore, we sought to determine whether it is possible to predict a given PHTS-affected individual's a priori risk of ASD, cancer, or the co-occurrence of both phenotypes. By integrating network proximity analysis performed on the human interactome, molecular simulations, and residue-interaction networks, we demonstrate the role of conformational dynamics in the structural communication and long-range allosteric regulation of germline PTEN variants associated with ASD or cancer. We show that the PTEN interactome shares significant overlap with the ASD and cancer interactomes, providing network-based evidence that PTEN is a crucial player in the biology of both disorders. Importantly, this finding suggests that a germline PTEN variant might perturb the ASD or cancer networks differently, thus favoring one disease outcome at any one time. Furthermore, protein-dynamic structural-network analysis reveals small-world structural communication mediated by highly conserved functional residues and potential allosteric regulation of PTEN. We identified a salient structural-communication pathway that extends across the inter-domain interface for cancer-only mutations. In contrast, the structural-communication pathway is predominantly restricted to the phosphatase domain for ASD-only mutations. Our integrative approach supports the prediction and potential modulation of the relevant conformational states that influence structural communication and long-range perturbations associated with mutational effects that lead to PTEN-ASD or PTEN-cancer phenotypes.
Keywords: PTEN; PTEN allostery; PTEN interactome; conformation dynamics; germline PTEN mutations; molecular dynamics simulation; network proximity analysis.
Copyright © 2019 American Society of Human Genetics. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Nelen M.R., van Staveren W.C., Peeters E.A., Hassel M.B., Gorlin R.J., Hamm H., Lindboe C.F., Fryns J.P., Sijmons R.H., Woods D.G. Germline mutations in the PTEN/MMAC1 gene in patients with Cowden disease. Hum. Mol. Genet. 1997;6:1383–1387. - PubMed
-
- Nelen M.R., Padberg G.W., Peeters E.A., Lin A.Y., van den Helm B., Frants R.R., Coulon V., Goldstein A.M., van Reen M.M., Easton D.F. Localization of the gene for Cowden disease to chromosome 10q22-23. Nat. Genet. 1996;13:114–116. - PubMed
-
- Liaw D., Marsh D.J., Li J., Dahia P.L., Wang S.I., Zheng Z., Bose S., Call K.M., Tsou H.C., Peacocke M. Germline mutations of the PTEN gene in Cowden disease, an inherited breast and thyroid cancer syndrome. Nat. Genet. 1997;16:64–67. - PubMed
-
- Eng C. PTEN: One gene, many syndromes. Hum. Mutat. 2003;22:183–198. - PubMed
-
- Tan M.H., Mester J., Peterson C., Yang Y., Chen J.L., Rybicki L.A., Milas K., Pederson H., Remzi B., Orloff M.S., Eng C. A clinical scoring system for selection of patients for PTEN mutation testing is proposed on the basis of a prospective study of 3042 probands. Am. J. Hum. Genet. 2011;88:42–56. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
