

A Forward Chemical Genetic Screen Reveals Gut Microbiota Metabolites That Modulate Host Physiology
- PMID: 31006530
- PMCID: PMC6536006
- DOI: 10.1016/j.cell.2019.03.036
A Forward Chemical Genetic Screen Reveals Gut Microbiota Metabolites That Modulate Host Physiology
Abstract
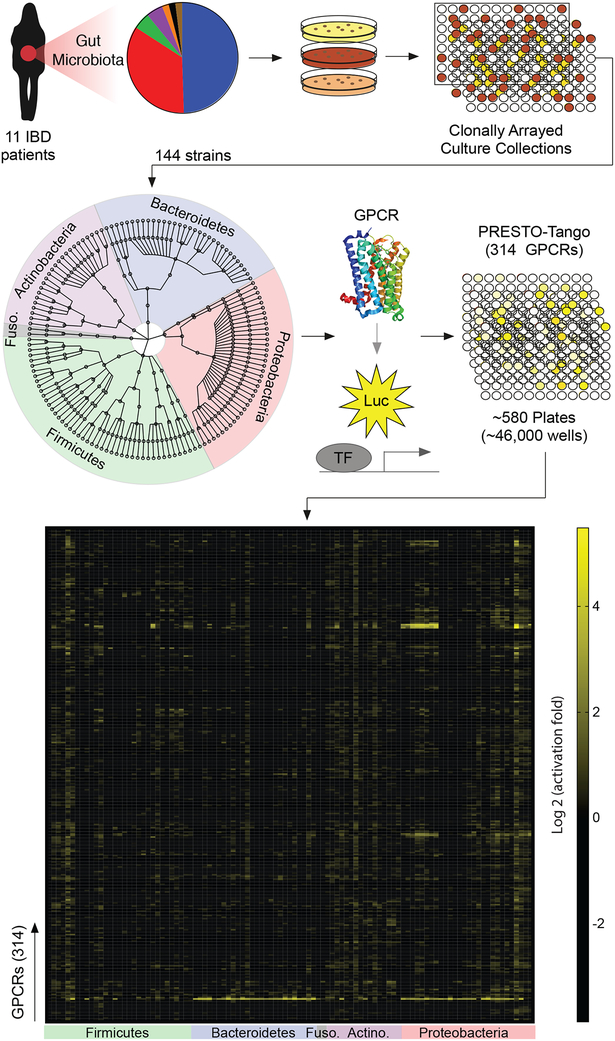
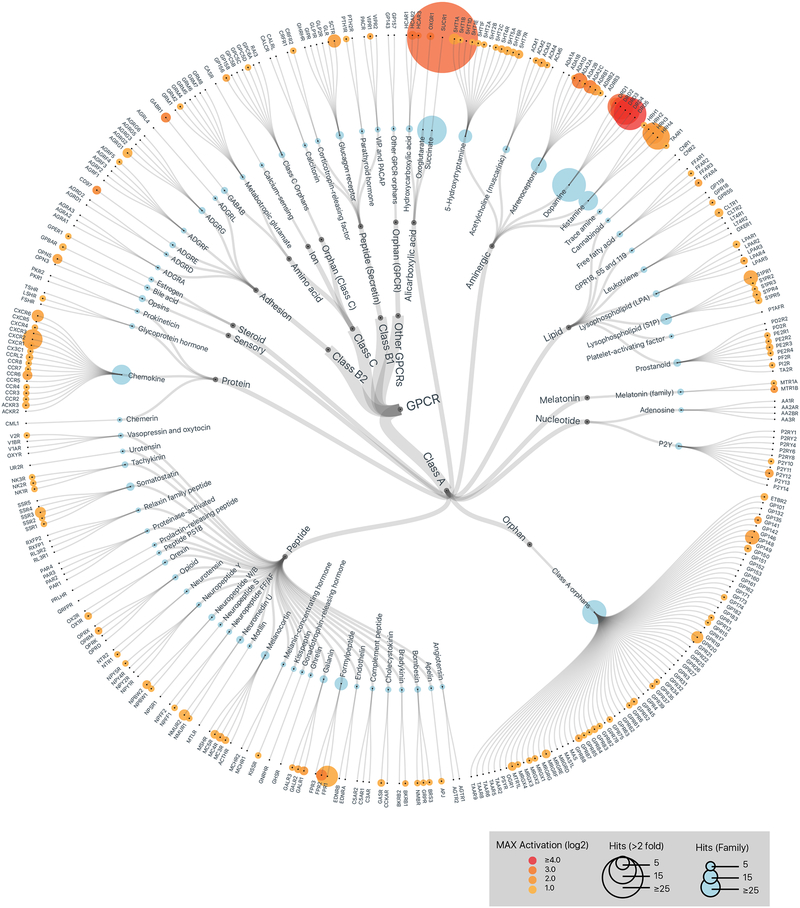
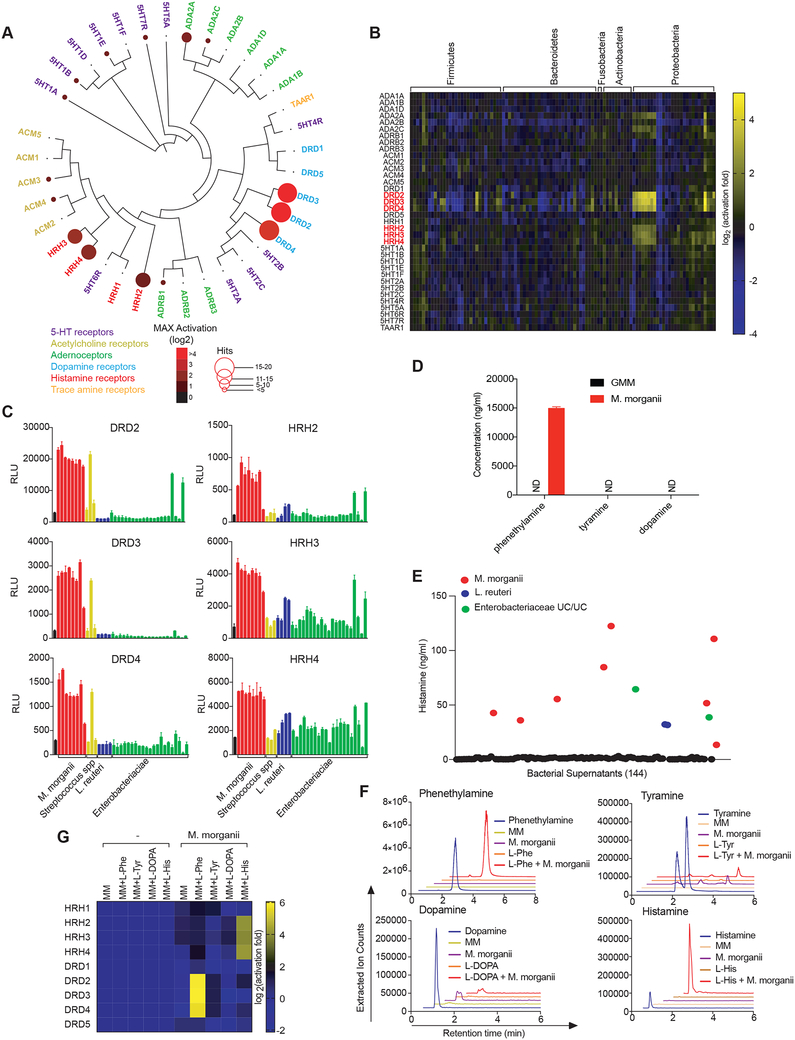
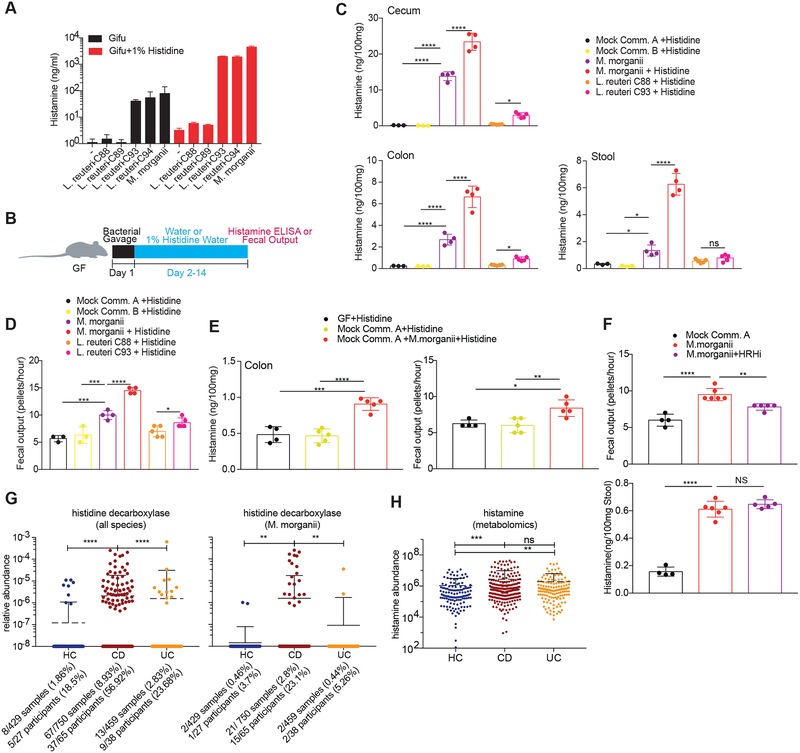
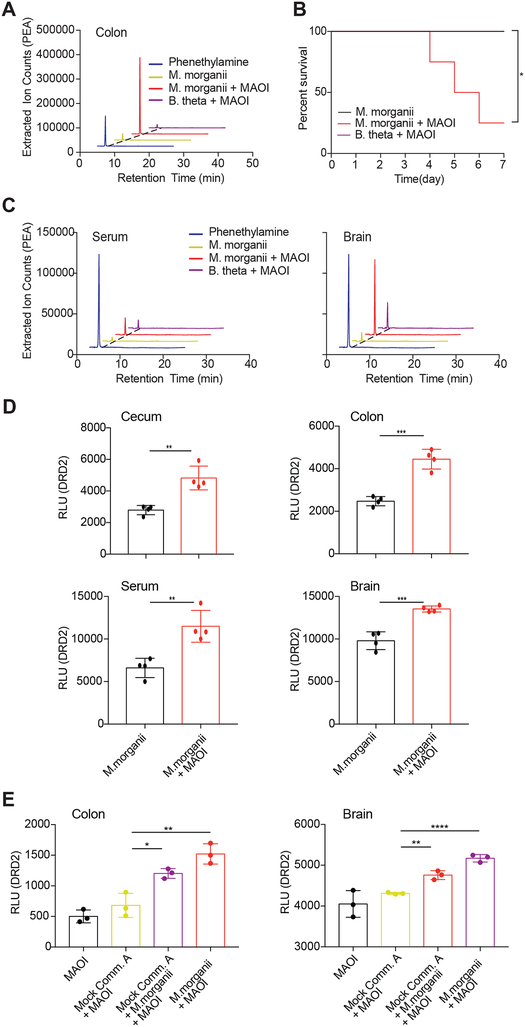
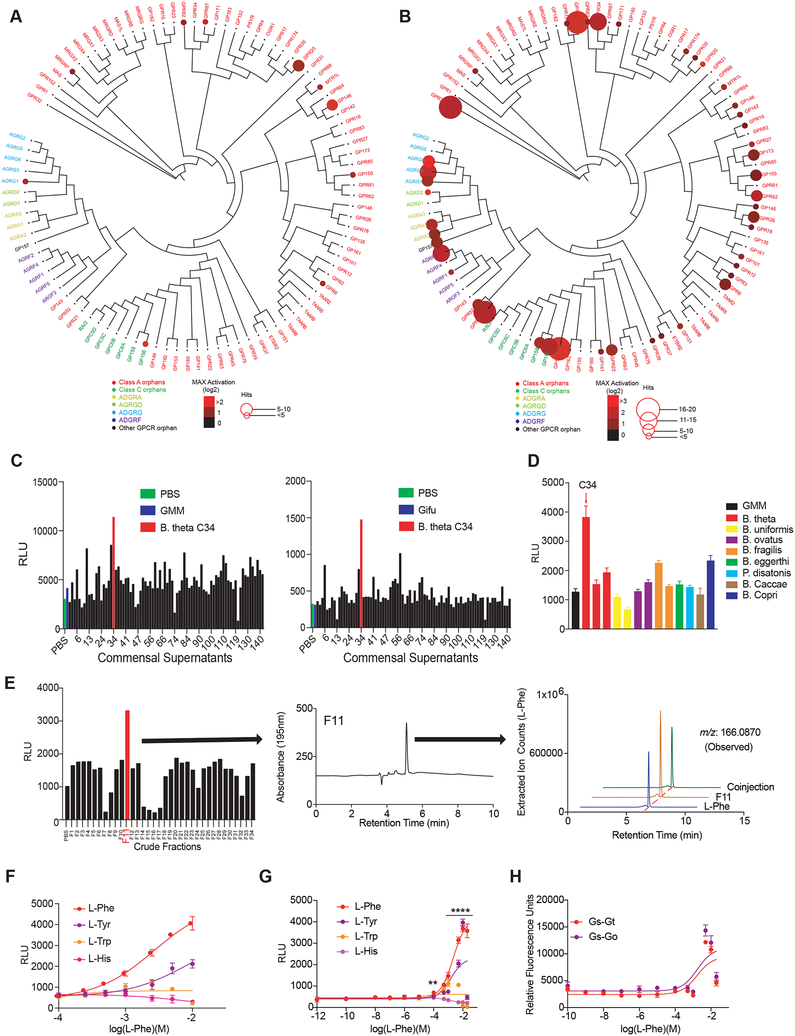
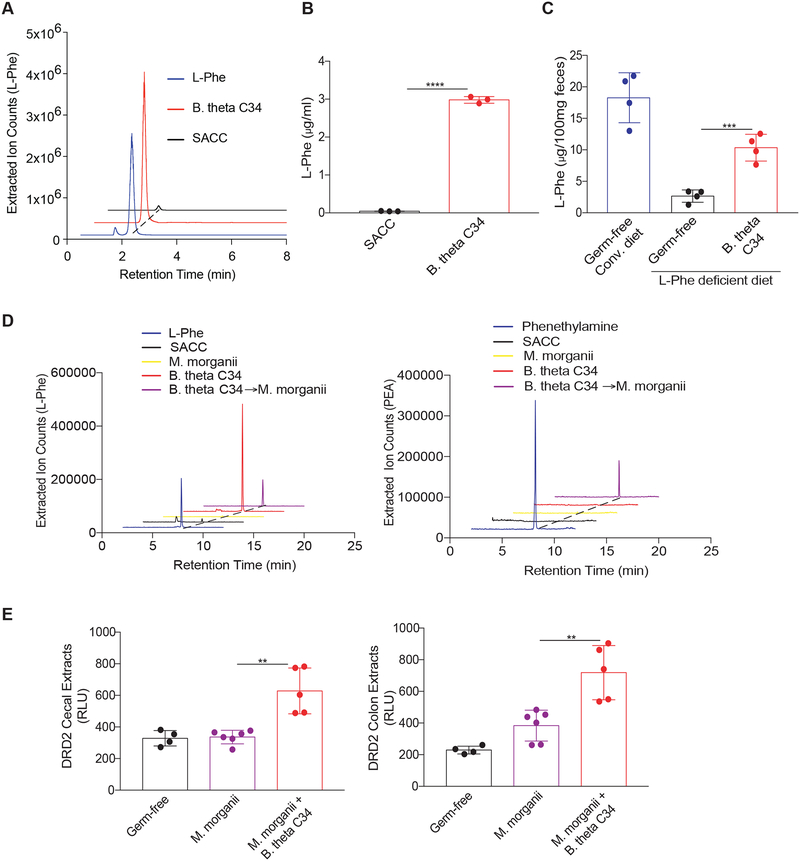
The intestinal microbiota produces tens of thousands of metabolites. Here, we used host sensing of small molecules by G-protein coupled receptors (GPCRs) as a lens to illuminate bioactive microbial metabolites that impact host physiology. We screened 144 human gut bacteria against the non-olfactory GPCRome and identified dozens of bacteria that activated both well-characterized and orphan GPCRs, including strains that converted dietary histidine into histamine and shaped colonic motility; a prolific producer of the essential amino acid L-Phe, which we identified as an agonist for GPR56 and GPR97; and a species that converted L-Phe into the potent psychoactive trace amine phenethylamine, which crosses the blood-brain barrier and triggers lethal phenethylamine poisoning after monoamine oxidase inhibitor administration. These studies establish an orthogonal approach for parsing the microbiota metabolome and uncover multiple biologically relevant host-microbiota metabolome interactions.
Copyright © 2019 Elsevier Inc. All rights reserved.
Figures
References
-
- Amisten S, Salehi A, Rorsman P, Jones PM, and Persaud SJ (2013). An atlas and functional analysis of G-protein coupled receptors in human islets of Langerhans. Pharmacol Ther 139, 359–391. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
