

Pyruvate-Carboxylase-Mediated Anaplerosis Promotes Antioxidant Capacity by Sustaining TCA Cycle and Redox Metabolism in Liver
- PMID: 31006591
- PMCID: PMC6585968
- DOI: 10.1016/j.cmet.2019.03.014
Pyruvate-Carboxylase-Mediated Anaplerosis Promotes Antioxidant Capacity by Sustaining TCA Cycle and Redox Metabolism in Liver
Abstract
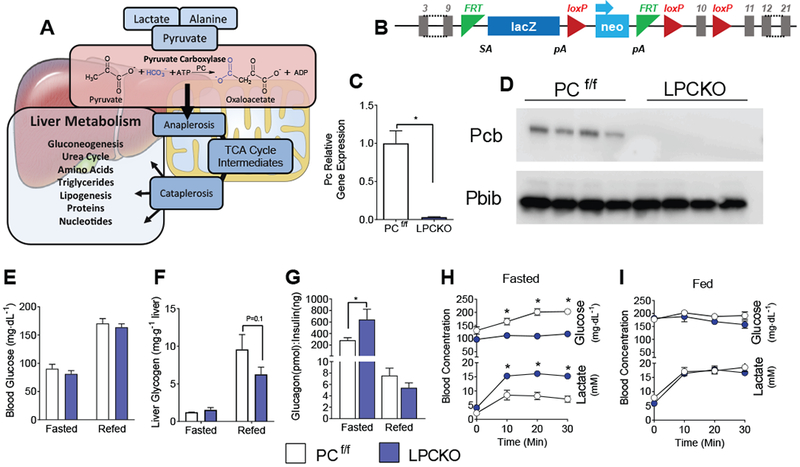
The hepatic TCA cycle supports oxidative and biosynthetic metabolism. This dual responsibility requires anaplerotic pathways, such as pyruvate carboxylase (PC), to generate TCA cycle intermediates necessary for biosynthesis without disrupting oxidative metabolism. Liver-specific PC knockout (LPCKO) mice were created to test the role of anaplerotic flux in liver metabolism. LPCKO mice have impaired hepatic anaplerosis, diminution of TCA cycle intermediates, suppressed gluconeogenesis, reduced TCA cycle flux, and a compensatory increase in ketogenesis and renal gluconeogenesis. Loss of PC depleted aspartate and compromised urea cycle function, causing elevated urea cycle intermediates and hyperammonemia. Loss of PC prevented diet-induced hyperglycemia and insulin resistance but depleted NADPH and glutathione, which exacerbated oxidative stress and correlated with elevated liver inflammation. Thus, despite catalyzing the synthesis of intermediates also produced by other anaplerotic pathways, PC is specifically necessary for maintaining oxidation, biosynthesis, and pathways distal to the TCA cycle, such as antioxidant defenses.
Keywords: TCA cycle; anaplerosis; gluconeogenesis; high-fat diet; liver physiology; metabolic flux; oxidative stress; pyruvate carboxylase; urea cycle.
Copyright © 2019 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests
The authors declare no competing interests.
Figures






Comment in
-
Pyruvate Carboxylase Wields a Double-Edged Metabolic Sword.Cell Metab. 2019 Jun 4;29(6):1236-1238. doi: 10.1016/j.cmet.2019.05.013. Cell Metab. 2019. PMID: 31167118
References
-
- Ahmad A, Kahler SG, Kishnani PS, Artigas-Lopez M, Pappu AS, Steiner R, Millington DS, and Van Hove JL (1999). Treatment of pyruvate carboxylase deficiency with high doses of citrate and aspartate. Am J Med Genet 87, 331–338. - PubMed
-
- Al-Sawaf O, Clarner T, Fragoulis A, Kan YW, Pufe T, Streetz K, and Wruck CJ (2015). Nrf2 in health and disease: current and future clinical implications. Clinical science (London, England : 1979) 129, 989–999. - PubMed
-
- Bigot A, Tchan MC, Thoreau B, Blasco H, and Maillot F (2017). Liver involvement in urea cycle disorders: a review of the literature. J Inherit Metab Dis 40, 757–769. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
