

Destabilization of chromosome structure by histone H3 lysine 27 methylation
- PMID: 31009462
- PMCID: PMC6510446
- DOI: 10.1371/journal.pgen.1008093
Destabilization of chromosome structure by histone H3 lysine 27 methylation
Abstract
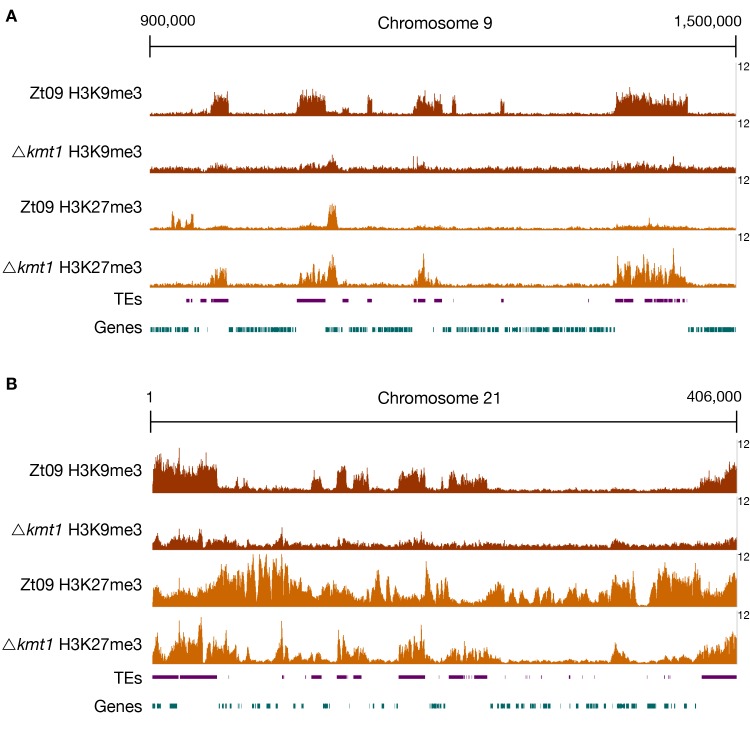
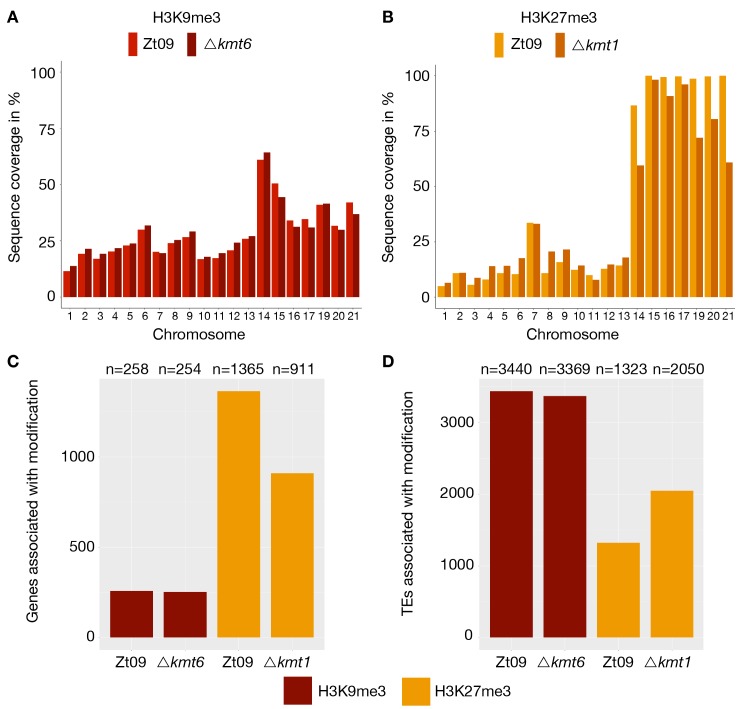
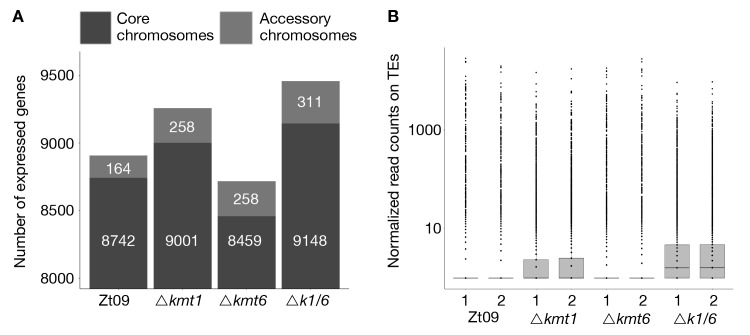
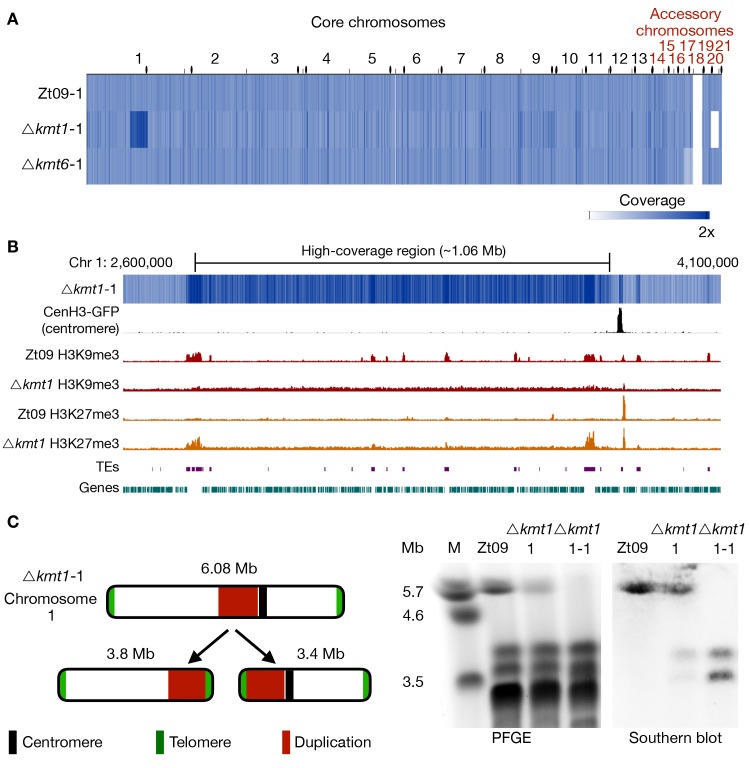
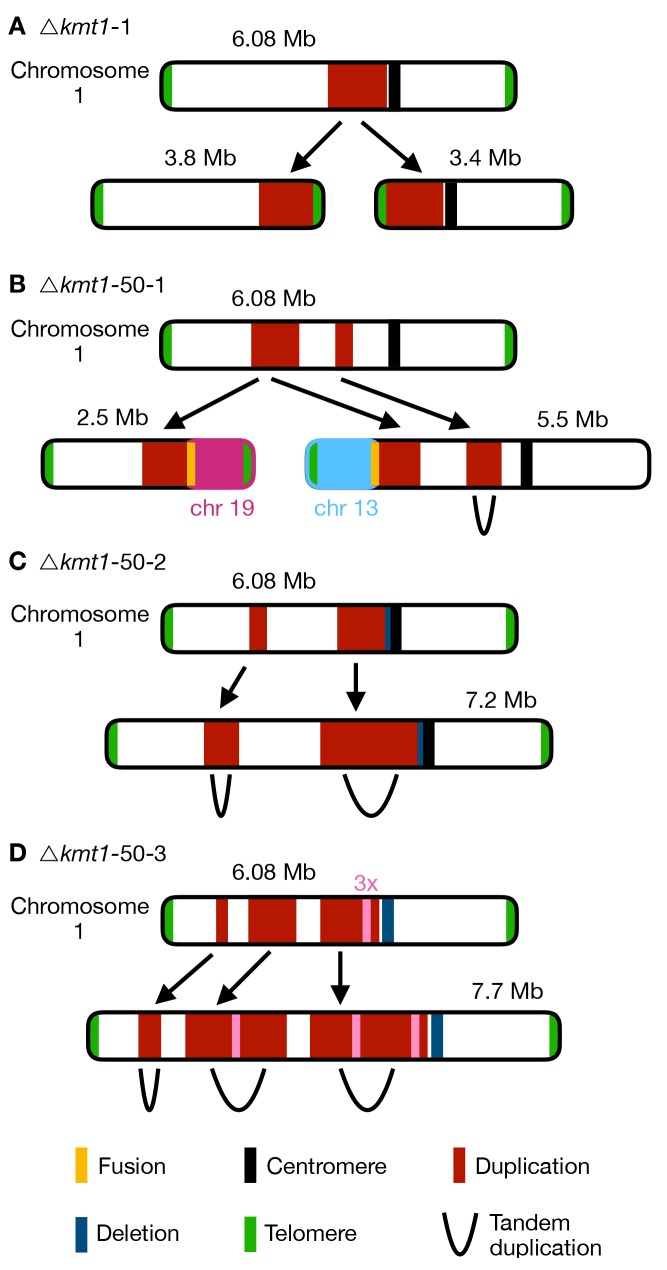
Chromosome and genome stability are important for normal cell function as instability often correlates with disease and dysfunction of DNA repair mechanisms. Many organisms maintain supernumerary or accessory chromosomes that deviate from standard chromosomes. The pathogenic fungus Zymoseptoria tritici has as many as eight accessory chromosomes, which are highly unstable during meiosis and mitosis, transcriptionally repressed, show enrichment of repetitive elements, and enrichment with heterochromatic histone methylation marks, e.g., trimethylation of H3 lysine 9 or lysine 27 (H3K9me3, H3K27me3). To elucidate the role of heterochromatin on genome stability in Z. tritici, we deleted the genes encoding the methyltransferases responsible for H3K9me3 and H3K27me3, kmt1 and kmt6, respectively, and generated a double mutant. We combined experimental evolution and genomic analyses to determine the impact of these deletions on chromosome and genome stability, both in vitro and in planta. We used whole genome sequencing, ChIP-seq, and RNA-seq to compare changes in genome and chromatin structure, and differences in gene expression between mutant and wildtype strains. Analyses of genome and ChIP-seq data in H3K9me3-deficient strains revealed dramatic chromatin reorganization, where H3K27me3 is mostly relocalized into regions that are enriched with H3K9me3 in wild type. Many genome rearrangements and formation of new chromosomes were found in the absence of H3K9me3, accompanied by activation of transposable elements. In stark contrast, loss of H3K27me3 actually increased the stability of accessory chromosomes under normal growth conditions in vitro, even without large scale changes in gene activity. We conclude that H3K9me3 is important for the maintenance of genome stability because it disallows H3K27me3 in regions considered constitutive heterochromatin. In this system, H3K27me3 reduces the overall stability of accessory chromosomes, generating a "metastable" state for these quasi-essential regions of the genome.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures


Similar articles
-
Histone H3 lysine 4 methyltransferase is required for facultative heterochromatin at specific loci.BMC Genomics. 2019 May 8;20(1):350. doi: 10.1186/s12864-019-5729-7. BMC Genomics. 2019. PMID: 31068130 Free PMC article.
-
H4K20me3 is important for Ash1-mediated H3K36me3 and transcriptional silencing in facultative heterochromatin in a fungal pathogen.PLoS Genet. 2023 Sep 25;19(9):e1010945. doi: 10.1371/journal.pgen.1010945. eCollection 2023 Sep. PLoS Genet. 2023. PMID: 37747878 Free PMC article.
-
Epigenetic plasticity safeguards heterochromatin configuration in mammals.Nucleic Acids Res. 2023 Jul 7;51(12):6190-6207. doi: 10.1093/nar/gkad387. Nucleic Acids Res. 2023. PMID: 37178005 Free PMC article.
-
Role of H3K9me3 heterochromatin in cell identity establishment and maintenance.Curr Opin Genet Dev. 2019 Apr;55:1-10. doi: 10.1016/j.gde.2019.04.013. Epub 2019 May 16. Curr Opin Genet Dev. 2019. PMID: 31103921 Free PMC article. Review.
-
Repressive Chromatin in Caenorhabditis elegans: Establishment, Composition, and Function.Genetics. 2018 Feb;208(2):491-511. doi: 10.1534/genetics.117.300386. Genetics. 2018. PMID: 29378810 Free PMC article. Review.
Cited by
-
Recent reactivation of a pathogenicity-associated transposable element is associated with major chromosomal rearrangements in a fungal wheat pathogen.Nucleic Acids Res. 2024 Feb 9;52(3):1226-1242. doi: 10.1093/nar/gkad1214. Nucleic Acids Res. 2024. PMID: 38142443 Free PMC article.
-
The contribution of DNA repair pathways to genome editing and evolution in filamentous pathogens.FEMS Microbiol Rev. 2022 Nov 2;46(6):fuac035. doi: 10.1093/femsre/fuac035. FEMS Microbiol Rev. 2022. PMID: 35810003 Free PMC article. Review.
-
Repeat-Induced Point Mutation and Gene Conversion Coinciding with Heterochromatin Shape the Genome of a Plant-Pathogenic Fungus.mBio. 2023 Jun 27;14(3):e0329022. doi: 10.1128/mbio.03290-22. Epub 2023 Apr 24. mBio. 2023. PMID: 37093087 Free PMC article.
-
Genome-wide mapping of histone modifications during axenic growth in two species of Leptosphaeria maculans showing contrasting genomic organization.Chromosome Res. 2021 Jun;29(2):219-236. doi: 10.1007/s10577-021-09658-1. Epub 2021 May 21. Chromosome Res. 2021. PMID: 34018080 Free PMC article.
-
Local Rather than Global H3K27me3 Dynamics Are Associated with Differential Gene Expression in Verticillium dahliae.mBio. 2021 Feb 22;13(1):e0356621. doi: 10.1128/mbio.03566-21. Epub 2022 Feb 8. mBio. 2021. PMID: 35130723 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
