

Novel Molecular Signatures in the PIP4K/PIP5K Family of Proteins Specific for Different Isozymes and Subfamilies Provide Important Insights into the Evolutionary Divergence of this Protein Family
- PMID: 31010098
- PMCID: PMC6523245
- DOI: 10.3390/genes10040312
Novel Molecular Signatures in the PIP4K/PIP5K Family of Proteins Specific for Different Isozymes and Subfamilies Provide Important Insights into the Evolutionary Divergence of this Protein Family
Abstract
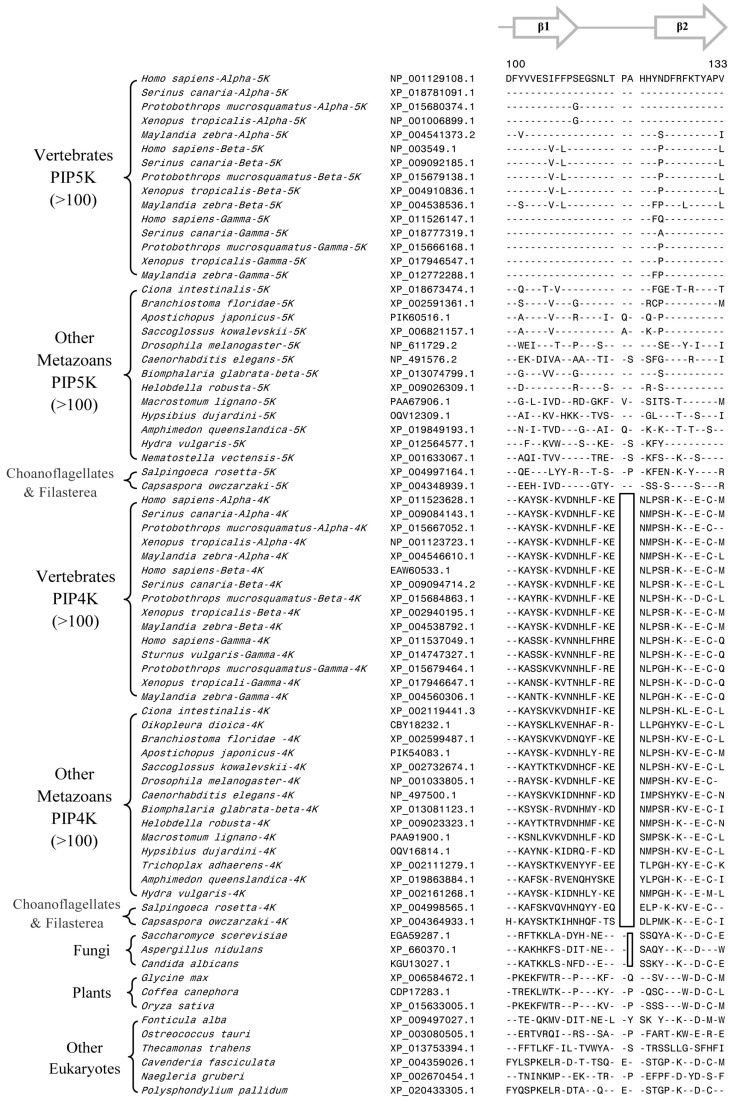
Members of the PIP4K/PIP5K family of proteins, which generate the highly important secondary messenger phosphatidylinositol-4,5-bisphosphate, play central roles in regulating diverse signaling pathways. In eukaryotic organisms, multiple isozymes and subfamilies of PIP4K/PIP5K proteins are found and it is of much interest to understand their evolution and species distribution and what unique molecular and biochemical characteristics distinguish specific isozymes and subfamilies of proteins. We report here the species distribution of different PIP4K/PIP5K family of proteins in eukaryotic organisms and phylogenetic analysis based on their protein sequences. Our results indicate that the distinct homologs of both PIP4K and PIP5K are found in different organisms belonging to the Holozoa clade of eukaryotes, which comprises of various metazoan phyla as well as their close unicellular relatives Choanoflagellates and Filasterea. In contrast, the deeper-branching eukaryotic lineages, as well as plants and fungi, contain only a single homolog of the PIP4K/PIP5K proteins. In parallel, our comparative analyses of PIP4K/PIP5K protein sequences have identified six highly-specific molecular markers consisting of conserved signature indels (CSIs) that are uniquely shared by either the PIP4K or PIP5K proteins, or both, or specific subfamilies of these proteins. Of these molecular markers, 2 CSIs are distinctive characteristics of all PIP4K homologs, 1 CSI distinguishes the PIP4K and PIP5K homologs from the Holozoa clade of species from the ancestral form of PIP4K/PIP5K found in deeper-branching eukaryotic lineages. The remaining three CSIs are specific for the PIP5Kα, PIP5Kβ, and PIP4Kγ subfamilies of proteins from vertebrate species. These molecular markers provide important means for distinguishing different PIP4K/PIP5K isozymes as well as some of their subfamilies. In addition, the distribution patterns of these markers in different isozymes provide important insights into the evolutionary divergence of PIP4K/PIP5K proteins. Our results support the view that the Holozoa clade of eukaryotic organisms shared a common ancestor exclusive of the other eukaryotic lineages and that the initial gene duplication event leading to the divergence of distinct types of PIP4K and PIP5K homologs occurred in a common ancestor of this clade. Based on the results gleaned from different studies presented here, a model for the evolutionary divergence of the PIP4K/PIP5K family of proteins is presented.
Keywords: conserved signature indels; evolution of the PIP4K/PIP5K family of proteins; holozoa clade of eukaryotic organisms; molecular signatures for the pip4k/pip5k isozymes and isoforms; phosphatidylinositol phosphate kinases; phylogenetic analysis; protein evolution; species distribution of pip4k/pip5k proteins.
Conflict of interest statement
The authors declare no conflict of interest.
Figures








Similar articles
-
Phylogenetic analysis of the diacylglycerol kinase family of proteins and identification of multiple highly-specific conserved inserts and deletions within the catalytic domain that are distinctive characteristics of different classes of DGK homologs.PLoS One. 2017 Aug 22;12(8):e0182758. doi: 10.1371/journal.pone.0182758. eCollection 2017. PLoS One. 2017. PMID: 28829789 Free PMC article.
-
Identification of a conserved 8 aa insert in the PIP5K protein in the Saccharomycetaceae family of fungi and the molecular dynamics simulations and structural analysis to investigate its potential functional role.Proteins. 2017 Aug;85(8):1454-1467. doi: 10.1002/prot.25306. Epub 2017 Apr 27. Proteins. 2017. PMID: 28407364
-
Origin and spread of photosynthesis based upon conserved sequence features in key bacteriochlorophyll biosynthesis proteins.Mol Biol Evol. 2012 Nov;29(11):3397-412. doi: 10.1093/molbev/mss145. Epub 2012 May 24. Mol Biol Evol. 2012. PMID: 22628531
-
Phylogeny and molecular signatures for the phylum Thermotogae and its subgroups.Antonie Van Leeuwenhoek. 2011 Jun;100(1):1-34. doi: 10.1007/s10482-011-9576-z. Epub 2011 Apr 19. Antonie Van Leeuwenhoek. 2011. PMID: 21503713 Review.
-
The phosphatidylinositol 4-phosphate 5-kinase family.Adv Enzyme Regul. 1996;36:115-40. doi: 10.1016/0065-2571(95)00005-4. Adv Enzyme Regul. 1996. PMID: 8869744 Review.
Cited by
-
Genomic Analyses Identify Novel Molecular Signatures Specific for the Caenorhabditis and other Nematode Taxa Providing Novel Means for Genetic and Biochemical Studies.Genes (Basel). 2019 Sep 24;10(10):739. doi: 10.3390/genes10100739. Genes (Basel). 2019. PMID: 31554175 Free PMC article.
-
A novel homeostatic mechanism tunes PI(4,5)P2-dependent signaling at the plasma membrane.J Cell Sci. 2023 Aug 15;136(16):jcs261494. doi: 10.1242/jcs.261494. Epub 2023 Aug 29. J Cell Sci. 2023. PMID: 37534432 Free PMC article.
-
Genome-Wide Identification of the PIP5K Gene Family in Camellia sinensis and Their Roles in Metabolic Regulation.Genes (Basel). 2024 Jul 17;15(7):932. doi: 10.3390/genes15070932. Genes (Basel). 2024. PMID: 39062711 Free PMC article.
-
Conserved Molecular Signatures in the Spike, Nucleocapsid, and Polymerase Proteins Specific for the Genus Betacoronavirus and Its Different Subgenera.Genes (Basel). 2022 Feb 25;13(3):423. doi: 10.3390/genes13030423. Genes (Basel). 2022. PMID: 35327976 Free PMC article.
-
Functional molecular evolution of a GTP sensing kinase: PI5P4Kβ.FEBS J. 2023 Sep;290(18):4419-4428. doi: 10.1111/febs.16763. Epub 2023 Mar 10. FEBS J. 2023. PMID: 36856076 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
